Признаки адаптации: Причины и признаки социальной адаптации
Причины и признаки социальной адаптации
Причины адаптации
Замечание 1
Социальная адаптация выступает в качестве очень сложного и многоуровневого процесса. Ее основная и ключевая причина – это стремление индивида приобщиться к социальным, культурным, экономическим и политическим условиям общества, в котором он ранее не был.
Желание познать что-то новое, получить новые знания, умения и навыки также считаются причинами социальной адаптации человека. На протяжение всего нашего существования мы постоянно меняем свои социальные роли, меняем статусы один на другой, стремимся облегчить свою жизнь, но вместе с тем сделать ее интереснее, насыщеннее. Каждому хочется жить в достатке, при этом имея крепкую семью, независимость и возможность самореализоваться. В связи с этим нам постоянно приходится сталкиваться с новыми социальными сферами, знакомиться с большим количеством людей, которые могут быть проводниками на новый жизненный этап, трансляторами ценностей и норм, правил поведения и традиций, которые царят в общественной группе.
Причины адаптации кроются также в нашем образовании. Жизнь делится на несколько этапов в плане профессионально-трудовой реализации. Изначально мы социализируемся дома, в семье. Благодаря родителям мы узнаем элементарные правила поведения, этикета, историю семьи, общества и нашего государства. Затем нам помещают в дошкольные заведения, где кроме родителей с нами работают педагоги. В детском саду мы постигаем азы коммуникаций, причем не только со взрослыми, но и со своими ровесниками. Таким образом, еще ребенком мы начинаем процессы адаптации к новым условиям: выходя за пределы семьи, мы сталкиваемся с ясельной группой, а также группой сверстников в детском саду.
Закончив этап социализации в дошкольных учреждениях, мы переходим на ступень выше – в школу. К ней мы адаптируемся, исходя из особенностей нашего интеллектуального, физического и психологического развития. На данном этапе с детьми работают психологи, которые помогают сглаживать негативные моменты адаптации: боязнь нового коллектива, стеснительность, отсутствие мотивации и активности. Причины нашей адаптации в школьных заведениях заключаются в том, что это – самый продолжительный этап обучения в одном и том же коллективе (9-11 лет), и одноклассники со временем должны восприниматься не просто как незнакомые посторонние люди, но и как семья.
Причины нашей адаптации в школьных заведениях заключаются в том, что это – самый продолжительный этап обучения в одном и том же коллективе (9-11 лет), и одноклассники со временем должны восприниматься не просто как незнакомые посторонние люди, но и как семья.
Оканчивая школу, следующим этапом идет поступление в вуз. Причина адаптации к вузовской среде – это комфорт пребывания в образовательном учреждении. Если человек чувствует себя уверенно, то его не будет беспокоить его неустроенность в коллективе. Эти мысли и негативные моменты не будут мешать ему заниматься самым главным – получать высшее образование, навыки и умения по выбранной профессии и специальности. Нередко говорят, что студенческая пора – самая прекрасная, поскольку человек реализует себя не только в образовательном процессе, но еще и во внеучебной деятельности: он принимает участие в организации важных мероприятий, становится частью общественных объединений. Его возраст и полученные навыки позволяют сформулировать свою общественную и гражданскую идею и продвигать ее в массы.
Причины, по которым индивид адаптируется к новым трудовым условиям, заключаются в необходимости реализации своего внутреннего капитала. Это значит, что человек осознает желание показать то, чему он обучался в течение получения высшего образования, желание продемонстрировать свои практические, эмпирические и творческие навыки.
Основные признаки адаптации
Адаптация, как и многие процессы, имеет свои признаки. Среди признаков успешной социальной адаптации выделяют следующие:
- Повышение уровня коммуникаций у индивида – прослеживается четкое желание идти на контакт со своими коллегами, руководством;
- Повышение трудоспособности и творческой активности – при успешной адаптации индивид не испытывает давления или чувства стеснения. Это помогает ему стать более работоспособным, не стесняться проявлять свою точку зрения и продвигать ее в массы;
- Позитивный настрой – хорошее настроение, медленная утомляемость;
- Желание помочь ближнему. Когда человек адаптируется в коллективе, он начинает видеть его как бы изнутри: все проблемы, перспективы, систему и сложную структуру.

Становятся видны и люди, которые недостаточно комфортно чувствуют себя, и индивид, осознавая то, что сам не так давно прошел через сложные процессы адаптации, проявляет инициативу и пытается помочь человеку в сложившейся ситуации. Это очень положительная интенция, поскольку, помогая ближнему, мы делимся с ним свои опытом, который зачастую бывает полезен.
Проблема дезадаптации
Нередко возникают ситуации, когда, по каким-то причинам, адаптация не происходит. Отсюда проистекает противоположный процесс – дезадаптация.
Определение 1
Дезадаптация- это психическое состояние индивида, при котором он частично или полностью утрачивает способность к адаптации к новым социально-культурным условиям.
Дезадаптация бывает нескольких основных видов:
- Патогенная дезадаптация;
- Психическая дезадаптация;
- Социальная дезадаптация.
Причины социальной дезадаптации скрываются в некоторых факторах. Например, в социальном (отторжение индивида самим социумом), социально-экономическом (недостаток материальных средств, низкий материальный статус, или наоборот – избыток материальных благ), психологическом (несовпадение установок индивида и социальной группы) или возрастном (отношения между детьми и родителями, между начальником и подчиненным).
Наиболее актуальный фактор, который порождает дезадаптацию – это социальный, и это признают очень многие исследователи. К этому фактору относят ошибки в процессе воспитания человека, нарушения в межличностных взаимосвязях субъекта и обширной социальной группы, ошибки в накоплении социального опыта (безработица, образование, не соответствующее интересам индивида). Все это ведет к достаточно сложным последствиям: индивид чувствует себя вне социума, не может определить свою роль, статус и принадлежность, теряет мотивацию и не осознает свою ценность в конкретной социальной, экономической или профессиональной группах.
АДАПТАЦИЯ | Энциклопедия Кругосвет
Содержание статьи- Генетическая основа.
- Питание.
- Защита.
- Размножение.
- Расселение.
АДАПТАЦИЯ, в биологии – развитие любого признака, который способствует выживанию вида и его размножению. Адаптации могут быть морфологическими, физиологическими или поведенческими.
Морфологические адаптации включают изменения формы или строения организма. Пример такой адаптации – твердый панцирь черепах, обеспечивающий защиту от хищных животных. Физиологические адаптации связаны с химическими процессами в организме. Так, запах цветка может служить для привлечения насекомых и тем самым способствовать опылению растения. Поведенческая адаптация связана с определенным аспектом жизнедеятельности животного. Типичный пример – зимний сон у медведя. Большинство адаптаций представляет собой сочетание перечисленных типов. Например, кровососание у комаров обеспечивается сложной комбинацией таких адаптаций, как развитие специализированных частей ротового аппарата, приспособленных к сосанию, формирование поискового поведения для нахождения животного-жертвы, а также выработка слюнными железами специальных секретов, которые предотвращают свертывание высасываемой крови.
Пример такой адаптации – твердый панцирь черепах, обеспечивающий защиту от хищных животных. Физиологические адаптации связаны с химическими процессами в организме. Так, запах цветка может служить для привлечения насекомых и тем самым способствовать опылению растения. Поведенческая адаптация связана с определенным аспектом жизнедеятельности животного. Типичный пример – зимний сон у медведя. Большинство адаптаций представляет собой сочетание перечисленных типов. Например, кровососание у комаров обеспечивается сложной комбинацией таких адаптаций, как развитие специализированных частей ротового аппарата, приспособленных к сосанию, формирование поискового поведения для нахождения животного-жертвы, а также выработка слюнными железами специальных секретов, которые предотвращают свертывание высасываемой крови.
Все растения и животные постоянно адаптируются к окружающей среде. Чтобы понять, как это происходит, необходимо рассматривать не только животное или растение в целом, но и генетическую основу адаптации.
Генетическая основа.
У каждого вида программа развития признаков заложена в генетическом материале. Материал и закодированная в нем программа передаются от одного поколения другому, оставаясь относительно неизменными, благодаря чему представители того или иного вида выглядят и ведут себя почти одинаково. Однако в популяции организмов любого вида всегда присутствуют небольшие изменения генетического материала и, следовательно, вариации признаков отдельных особей. Именно из этих разнообразных генетических вариаций процесс приспособления отбирает те признаки или благоприятствует развитию таких признаков, которые в наибольшей степени увеличивают шансы на выживание и тем самым на сохранение генетического материала. Адаптация, таким образом, может рассматриваться как процесс, посредством которого генетический материал повышает свои шансы на сохранение в последующих поколениях. С этой точки зрения, каждый вид олицетворяет собой успешный способ сохранения определенного генетического материала.
Чтобы передать генетический материал, особь любого вида должна иметь возможность питаться, дожить до периода размножения, оставить потомство и затем распространить его на возможно большей территории.
Питание.
Все растения и животные должны получать из окружающей среды энергию и различные вещества, прежде всего кислород, воду и неорганические соединения. Почти все растения используют энергию Солнца, трансформируя ее в процессе фотосинтеза. Животные получают энергию, питаясь растениями или другими животными.
Каждый вид определенным образом приспособлен к тому, чтобы обеспечивать себя питанием. Ястребы имеют острые когти для захватывания добычи, а расположение глаз в передней части головы позволяет им оценить глубину пространства, что необходимо для охоты при полете на большой скорости. У других птиц, например цапель, развились длинные шея и ноги. Они добывают пищу, осторожно бродя по мелководью и подстерегая зазевавшихся водных животных. Дарвиновы вьюрки – группа близкородственных видов птиц с Галапагосских островов – представляют классический пример высокоспециализированной адаптации к разным способам питания. Благодаря тем или иным адаптивным морфологическим изменениям, в первую очередь в строении клюва, одни виды стали зерноядными, другие – насекомоядными.
Благодаря тем или иным адаптивным морфологическим изменениям, в первую очередь в строении клюва, одни виды стали зерноядными, другие – насекомоядными.
Если обратиться к рыбам, то хищники, например акулы и барракуды, имеют острые зубы для поимки добычи. Другие, например мелкие анчоусы и сельди, добывают мелкие частицы пищи путем фильтрации морской воды через гребневидные жаберные тычинки.
У млекопитающих прекрасным примером адаптации к типу питания служат особенности строения зубов. Клыки и коренные зубы у леопардов и других кошачьих исключительно остры, что позволяет этим животным удерживать и разрывать тело жертвы. У оленей, лошадей, антилоп и других пастбищных животных большие коренные зубы имеют широкие ребристые поверхности, приспособленные для пережевывания травы и иной растительной пищи.
Разнообразные способы получения питательных веществ можно наблюдать не только у животных, но и у растений. Многие из них, в первую очередь бобовые – горох, клевер и другие – развили симбиотические, т. е. взаимовыгодные, отношения с бактериями: бактерии переводят атмосферный азот в химическую форму, доступную для растений, а растения предоставляют бактериям энергию. Насекомоядные растения, такие, как саррацения и росянка, получают азот из тел насекомых, пойманных ловчими листьями.
е. взаимовыгодные, отношения с бактериями: бактерии переводят атмосферный азот в химическую форму, доступную для растений, а растения предоставляют бактериям энергию. Насекомоядные растения, такие, как саррацения и росянка, получают азот из тел насекомых, пойманных ловчими листьями.
Защита.
Окружающая среда состоит из живых и неживых компонентов. Живое окружение любого вида включает животных, питающихся особями этого вида. Адаптации хищных видов направлены на эффективную добычу пищи; виды-жертвы приспосабливаются, чтобы не стать добычей хищников.
Многие виды – потенциальные жертвы – имеют защитную или маскирующую окраску, которая скрывает их от хищников. Так, у некоторых видов оленей пятнистая шкура молодых особей незаметна на фоне чередующихся пятен света и тени, а зайцев-беляков трудно различить на фоне снежного покрова. Длинные тонкие тела насекомых-палочников тоже трудно увидеть, потому что они напоминают сучки или веточки кустов и деревьев.
У оленей, зайцев, кенгуру и многих других животных развились длинные ноги, позволяющие им убегать от хищников. Некоторые животные, например опоссумы и свиномордые ужи, даже выработали своеобразный способ поведения – имитацию смерти, которая повышает их шансы на выживание, поскольку многие хищники не едят падали.
Некоторые животные, например опоссумы и свиномордые ужи, даже выработали своеобразный способ поведения – имитацию смерти, которая повышает их шансы на выживание, поскольку многие хищники не едят падали.
Некоторые виды растений покрыты шипами или колючками, отпугивающими животных. Многие растения имеют отвратительный для животных вкус.
Факторы окружающей среды, в частности климатические, нередко ставят живые организмы в трудные условия. Например, животным и растениям часто приходится приспосабливаться к крайним значениям температуры. Животные спасаются от холода, используя изолирующий мех или перья, мигрируя в места с более теплым климатом или впадая в зимнюю спячку. Большинство растений переживает холода, переходя в состояние покоя, эквивалентное спячке у животных.
В жару охлаждение животного происходит за счет потоотделения или частого дыхания, увеличивающего испарение. Некоторые животные, в особенности пресмыкающиеся и земноводные, способны впадать в летнюю спячку, которая по сути аналогична зимней, но вызвана жарой, а не холодом. Другие просто ищут прохладное место.
Другие просто ищут прохладное место.
Растения могут до некоторой степени поддерживать свою температуру, регулируя интенсивность испарения, которое имеет то же охлаждающее действие, что и потоотделение у животных.
Размножение.
Критическим этапом в обеспечении непрерывности жизни является размножение – процесс, в ходе которого происходит передача генетического материала следующему поколению. Размножение имеет два важных аспекта: встречу разнополых особей для обмена генетическим материалом и выращивание потомства.
К числу адаптаций, обеспечивающих встречу особей разного пола, относится звуковая коммуникация. У некоторых видов большую роль в этом смысле играет обоняние. Например, котов сильно привлекает запах кошки в период течки. Многие насекомые выделяют т.н. аттрактанты – химические вещества, привлекающие особей противоположного пола. Запахи цветков являются эффективной адаптацией растений для привлечения насекомых-опылителей. Некоторые цветки сладко пахнут и привлекают питающихся нектаром пчел; другие пахнут отвратительно, привлекая мух, питающихся на падали.
Зрение тоже очень важно для встречи особей разного пола. У птиц брачное поведение самца, его пышные перья и яркая окраска привлекают самку и подготавливают ее к копуляции. Окраска цветка у растений часто указывает, какое животное необходимо для опыления этого растения. Например, цветки, опыляемые колибри, окрашены в красный цвет, который привлекает этих птиц.
Многие животные выработали способы защиты своего потомства в начальный период жизни. Большинство адаптаций такого рода относятся к поведенческим и включают такие действия одного или обоих родителей, которые повышают шансы на выживание детенышей. Большинство птиц строит гнезда, характерные для каждого вида. Однако некоторые виды, например воловья птица, откладывают яйца в гнезда других видов птиц и вверяют детенышей родительской заботе вида-хозяина. У многих птиц и млекопитающих, а также у некоторых рыб имеется период, когда один из родителей идет на большой риск, беря на себя функцию защиты потомства. Хотя такое поведение иногда грозит гибелью родителю, оно обеспечивает безопасность потомства и сохранение генетического материала.
Целый ряд видов животных и растений использует иную стратегию размножения: они производят на свет огромное число потомков и оставляют их незащищенными. В этом случае низкие шансы на выживание у отдельной подрастающей особи оказываются сбалансированы многочисленностью потомства. См. также РАЗМНОЖЕНИЕ.
Расселение.
Большинство видов выработало механизмы для удаления потомства от тех мест, где оно появилось на свет. Этот процесс, называемый расселением, увеличивает вероятность того, что потомство будет подрастать на еще не занятой территории.
Большинство животных просто избегает мест, где слишком сильна конкуренция. Однако накапливаются свидетельства в пользу того, что расселение обусловлено генетическими механизмами.
Многие растения приспособились к распространению семян с помощью животных. Так, соплодия дурнишника имеют на поверхности крючочки, которыми они цепляются за шерсть проходящих мимо животных. Другие растения образуют вкусные мясистые плоды, например ягоды, которые поедаются животными; семена проходят через пищеварительный тракт и неповрежденными «высеваются» в другом месте. Для распространения растения используют и ветер. Например, ветром переносятся «пропеллеры» семян клена, а также семена ваточника, имеющие хохолки из тонких волосков. Степные растения типа перекати-поле, приобретающие к моменту созревания семян шарообразную форму, перегоняются ветром на большие расстояния, по пути рассеивая семена.
Для распространения растения используют и ветер. Например, ветром переносятся «пропеллеры» семян клена, а также семена ваточника, имеющие хохолки из тонких волосков. Степные растения типа перекати-поле, приобретающие к моменту созревания семян шарообразную форму, перегоняются ветром на большие расстояния, по пути рассеивая семена.
Выше были приведены лишь некоторые наиболее яркие примеры адаптаций. Однако практически каждый признак любого вида является результатом адаптации. Все эти признаки составляют гармоничную совокупность, что позволяет организму успешно вести свой особый образ жизни. Человек во всех его признаках, от структуры головного мозга до формы большого пальца на ноге, является результатом адаптации. Адаптивные признаки способствовали выживанию и размножению его предков, имевших те же самые признаки. В целом концепция адаптации имеет большое значение для всех направлений биологии. См. также НАСЛЕДСТВЕННОСТЬ.
Что такое социальная адаптация, какие уровни и этапы адаптации бывают?
У каждого из нас в процессе эволюции выработался механизм привыкания и приспособления.
Человек — существо социальное, поэтому, попадая в различные группы, общества и условия, он начинает приспосабливаться и взаимодействовать с новым окружением. Это называется «адаптация в обществе».
Из этой статьи вы узнаете, что такое социально-психологическая адаптация, какие существуют признаки адаптации в социуме и как понять, что человек социально не адаптирован.
Что такое социальная адаптация
Слово «адаптация» в переводе с латинского «adaptatio» значит «приспособление», «приноровление».
Понятие адаптации изначально использовалось в биологии и означало способность человеческого организма реагировать и изменяться под воздействием окружающей среды.
Что касается социологии, то термину «адаптация» дали такое определение — это процесс приспособления одного индивида или целой группы к различным социальным группам, средам, способность человека отвечать ее требованиям и условиям.
Кратко процесс приспособления можно расписать в виде схемы: конфликт — тревога — защитная реакция и приспособление.
Соц адаптация происходит за счет изменения поведения, обучения, «прокачки» навыков, смены установок, потребления информации, принятия тех или иных решений, анализа результатов.
Виды и уровни социальной адаптации
В психологии показателем успешной адаптации является включение личности в основные процессы какой-либо социальной среды и успешное освоение принятых правил и норм. Социальной средой может выступать семья, рабочий коллектив, группа учащихся и т.д.
Специалисты разделяют социальное приспособление на несколько видов, в зависимости от целей, особенностей новой среды и других факторов:
- психологическая — приспособление человека в обществе путем выстраивания психологических связей и стимулов для освоения различных психологических и социальных ролей
-
профессиональная — освоение, привыкание, принятие правил, норм, традиций в новой профессии и трудовом коллективе.
 А также приспособление к новым условиям труда
А также приспособление к новым условиям труда - педагогическая — процесс получения индивидом образования, освоения навыков и воспитание. В качестве примера социальной и педагогической адаптации можно привести нахождение ребенка в детском саду (освоение необходимых навыков и развитие коммуникации для поступления в школу), обучение в школе, колледже, институте и т.д.
- экономическая — люди приспосабливаются к новым для них нормам социально-экономического характера.
В психологии выделяют несколько уровней адаптации, в зависимости т того, насколько сильно в человеке проявлена способность к адаптивности (приспособлению):
|
Уровни приспособления |
Сущность и характеристика |
|
Низкий уровень |
Этот уровень характеризует низкая адаптированность индивида. |
|
Средний уровень |
Этому типу характерна нормальная адаптация индивида, но при этом, если уровень приспособления для той или иной серы недостаточное, то человек не будет испытывать дискомфорта и психологических проблем. |
|
Высокий (нормальный) уровень |
Человек адаптирован по всем направлениям, у него стабильный психоэмоциональный фон. |
|
Избыточный |
Уровень приспособления настолько велик, что человек испытывает дискомфорт, стресс и становится подвержен психоэмоциональным расстройствам. |
 По-другому, этот уровень называют «дезадаптация». Социально неадаптированный человек не может приспособиться к новым ролям и изменяемым условиям, он склонен к депрессиям, психоэмоциональным расстройствам, паническим атакам и фобиям.
По-другому, этот уровень называют «дезадаптация». Социально неадаптированный человек не может приспособиться к новым ролям и изменяемым условиям, он склонен к депрессиям, психоэмоциональным расстройствам, паническим атакам и фобиям.

Уровень социальной адаптации означает способность человека «вживаться» в непривычную для себя роль и осваивать механизмы социальной адаптации с минимальными негативными последствиями для собственного здоровья.
Пример
Ольга устроилась на новую работу в международную компанию, переход на новую должность в новый офис совпал со временем выхода из декретного отпуска. Несколько месяцев Ольга не могла привыкнуть к новому распорядку дня, испытывала страх перед поездкой на работу, сомневалась в себе, что не справится с новыми обязанностями, тревожилась по каждому даже самому простому вопросу. Понимая, что этот дисбаланс мешает ей работать и жить нормальной жизнью, Ольга обратилась к психологу. Специалист сразу сказал, что у Ольги проблемы с адаптацией. На консультации он определил неблагоприятные факторы, дал советы, как восстановить эмоциональный баланс и убрать проблемы с привыканием к новой работе.
Симптомы дезадаптации
Симптомы дезадаптации проявляются у каждого человека по разному, они могут затрагивать как физиологическую или психологическую сферу. Вот наиболее часто встречающиеся симптомы:
- быстрая утомляемость, повышенная раздражительность
- нарушение пищевых привычек и расстройство сна
- повышенная возбудимость, чрезмерное эмоциональное реагирование на любой раздражитель
- повышенная тревожность, частая перемена настроения
- тошнота, головокружение
- мышечные спазмы, резкое изменение давления, пульса, затрудненное дыхание
- снижение концентрации, продуктивности, внимательности
- безразличие к работе, коллективу, окружающим, игнорирование окружающих
Тяжелая форма дезадаптации может сопровождаться депрессивным настроением, суицидальными мыслями, самоповреждающим поведением.
Запишитесь на онлайн-консультацию, если вы обнаружили у себя хотя бы один симптом дезадаптации. Наши психологи по телефону расскажут вам о специальных программах по работе с нервно-психическими расстройствами, помогут вам справиться с дезадаптацией и вернуть уверенность в собственных силах.
Этапы социальной адаптации детей и подростков
Процесс социального приспособления начинается еще в младенчестве и активно продолжается до подросткового возраста.
Специалисты разделяют особенности социальной адаптации с новой средой по следующим возрастам:
- первый год жизни ребенка — младенец полностью зависит от родителей и взаимодействует только с ними
- от 1 года до 3 лет — ребенок начинает взаимодействовать с другими членами семьи, бессознательное копирование поведения родителей
- от 3 до 6 лет — приспособление к другим детям в детском саду, сознательное копирование поведения родителей и других членов семьи
- от 6 до 10 лет — привыкание к школе и во время учебы в школе, взаимодействие с одноклассниками, активное изучение новых ролей
- от 10 до 15 лет — период полового созревания, активное взаимодействие с окружающими, эмоциональный всплеск, активное проявление чувств и изучение своих желаний и потребностей, попытки самореализации
Важно! Детско-родительские отношения напрямую влияют на процесс адаптации ребенка. На родителях лежит важнейшая задача по передаче ребенку навыков взаимодействия с окружающими — ошибки в воспитании могут привести негативным последствиям.
На родителях лежит важнейшая задача по передаче ребенку навыков взаимодействия с окружающими — ошибки в воспитании могут привести негативным последствиям.
Основные стадии адаптации
Любой человек в процессе приспособления проходит через 4 стадии:
- Стадия когнитивного диссонанса — когда личность не осознает и не принимает новые нормы и правила поведения. Происходит знакомство с новой средой и новыми ролями.
- Индивид признает существование новой среды
- Индивид начинает процесс приспособления — соблюдает правила, ценности, установки
- Этап полной адаптации индивида к новой среде — изменяется поведение, ощущение, происходит взаимное принятие друг друга (индивида и среды).
Запишитесь на онлайн-консультацию, если у вас или у вашего ребенка возникли проблемы с адаптацией. Наши психологи проведут индивидуальную беседу с вами или с вашим чадом и помогут безболезненно пройти все этапы социальной адаптации.
Частые вопросы
Что такое социально-психологическая адаптация ?
+
Это процесс приспособления человека или группы людей к новым условиям жизни, к новым общественным связям, обучение взаимодействию с окружающими и принятие новых правил.
Какие функции влияют на социальное привыкание?
+
Процесс приспособления зависит от личностных качеств человека его темперамента, активности, жизненной позиции, зрелости психической системы, способности к обучению, жизненных целей, установок и ориентиров.
Причины дезадаптации?
+
Чтобы дать точный ответ на этот вопрос, нужно знать, какие стрессовые факторы «запустили» процесс дезадаптации. Специалисты выделяют следующие негативные моменты: насилие, сильный стресс, расстройства нервной системы (как врожденные, так и приобретенные), психосоматические проблемы и др.
Как помочь ребенку адаптироваться к социуму?
+
Лучше всего это делать через личный пример — знакомьтесь с новыми людьми, налаживайте новые контакты, учите самостоятельности, не критикуйте и позволяйте ребенку самому анализировать негативные моменты и ситуации.
Заключение эксперта
За всю свою жизнь человек много раз меняет свои социальные роли. Начиная с самого детства он вступает в тесную взаимосвязь с родителями, членами семьи, окружающими. Если на каком-то этапе возникают проблемы с приспособлением к новым правилам иной социальной группы, то индивид испытывает стресс, становится нервным, вспыльчивым, агрессивным или, наоборот, старается избегать любых контактов и закрывается эмоционально.
Главная функция социальной адаптации — формирование человека, как личности, налаживание социального взаимодействия, комфортное приспособление индивида к меняющимся условиям.
Публикуем только проверенную информацию
Автор статьи
Монахова Альбина Петровна клинический психолог
Стаж 17 лет
Консультаций 1439
Статей 226
Специалист в области клинической психологии. Помощь в поиске инструментов для самореализации, проработка убеждений, страхов и тревог. Работа с самоотношением, внутренними границами, понимание взаимодействия с социумом через осознанные личностные изменения.
- 2007 — 2008 год МУЗ Детская поликлиника №4 — педагог психолог
- 2008 — 2009 ООО Здоровая страна — клинический психолог
- 2009 — 2021 год Республиканский наркологический диспансер — психолог
- 2012 — 2013 год Профессиональная медицина — психолог
- 2013 — 2015 год ООО Возрождение — психолог
- 2019 по настоящее время ООО Теледоктор24 — психолог
Особенности адаптации первоклассников
Первый учебный год в школе — переломный период, когда меняется привычный уклад жизни, меняется место ребенка в системе общественных отношений, появляются новые обязанности, увеличивается нагрузка на детскую психику. Беззаботное дошкольное время сменяется ежедневной работой — учебными занятиями, которые требуют от первоклассника напряженного умственного труда, активизации внимания, усидчивости и правильной рабочей позы в течение всего урока.
Первый раз в первый класс: дети в первом классе, или как ребенок привыкает к школе.

Ребенку нужно установить контакты с одноклассниками и учителями, научиться выполнять новые обязанности, новые требования дисциплины. Первые недели учебы, а иногда и месяцы, организму ребенка трудно привыкнуть к новым требованиям и ритму жизни. Процесс адаптации можно разделить на несколько общепринятых этапов:
Первый этап– ориентировочный, характеризующийся бурной реакцией и значительным напряжением практически всех систем организма. Длится 2-3 недели.
Второй этап– неустойчивое приспособление, когда организм ищет и находит какие-то оптимальные варианты реакций на эти воздействия. На втором этапе затраты снижается, бурная реакция начинает затихать.
Третий этап– период относительно устойчивого приспособления, когда организм находит наиболее подходящие варианты реагирования на нагрузку, требующие меньшего напряжения всех систем. В период адаптации первоклассники могут жаловаться на усталость, общее недомогание, наблюдается плаксивость, раздражительность. Нередко нарушается сон, снижается аппетит. У детей в начальных классах могут возникнуть трудности психологического характера: страх перед учителем, отрицательное отношение к учебе, недооценка своих способностей и возможностей. Этот период адаптации ребенка к школе называют иногда «школьным стрессом». Кроме того, что ребенок в 7 лет идет в школу и испытывает стресс, связанный с адаптацией к новым условиям, в этом же возрасте отмечается очередной возрастной кризис, который сопровождается изменениями в организме ребенка: быстрый рост, изменения в работе нервной, сердечно-сосудистой, дыхательной и других систем. В период возрастного кризиса у ребенка происходят изменения и в характере. Родителям нужно более внимательно относиться к детям в этот переломный для них момент. Важно замечать все изменения в детском поведении, пытаться понять, чем они обусловлены, помочь направить ребенка в нужное русло. Возможно обращение к врачам – специалистам.
Нередко нарушается сон, снижается аппетит. У детей в начальных классах могут возникнуть трудности психологического характера: страх перед учителем, отрицательное отношение к учебе, недооценка своих способностей и возможностей. Этот период адаптации ребенка к школе называют иногда «школьным стрессом». Кроме того, что ребенок в 7 лет идет в школу и испытывает стресс, связанный с адаптацией к новым условиям, в этом же возрасте отмечается очередной возрастной кризис, который сопровождается изменениями в организме ребенка: быстрый рост, изменения в работе нервной, сердечно-сосудистой, дыхательной и других систем. В период возрастного кризиса у ребенка происходят изменения и в характере. Родителям нужно более внимательно относиться к детям в этот переломный для них момент. Важно замечать все изменения в детском поведении, пытаться понять, чем они обусловлены, помочь направить ребенка в нужное русло. Возможно обращение к врачам – специалистам.
Адаптация ребенка к школе — процесс многоплановый. Его составляющими являются социально-психологическая и физиологическая адаптации. Эти составляющие связаны между собой.
Его составляющими являются социально-психологическая и физиологическая адаптации. Эти составляющие связаны между собой.
Социально-психологическая адаптация
Когда ребенок впервые идет в школу, у него появляется новая социальная роль — роль ученика. Изменение социального статуса влечет за собой изменение самосознания первоклассника. То, что было значимым в дошкольном возрасте, становится второстепенным, а более ценным становится то, что имеет отношение к учебе. Помочь плавно подняться на ступень выше (от дошкольника к школьнику) помогут учитель и родители ребенка. Педагог поможет детям познакомиться друг с другом, с учителями, с правилами школы. Родители создадут комфортную обстановку дома. Некоторые первоклашки, уходя из дома в школу, берут с собой игрушки. Не стоит запрещать этого делать. Рядом с любимой куклой или машинкой ребенок чувствует себя защищенным. Игрушка — это частичка любимого и безопасного дома. Особенно важно чувство защищенности для робких и застенчивых детей. У детей совершенно по – разному проходит социально-психологическая адаптация:
У детей совершенно по – разному проходит социально-психологическая адаптация:
Адаптация 1,5-2 месяца. Такие дети быстро устанавливают контакт с одноклассниками и учителями, легко осваиваются в школе, заводят новых друзей. Они без особых усилий выполняют требования учителя, всегда в хорошем настроении, добросовестно выполняют задания. Такие дети обычно легко преодолевают возникающие трудности. Дети быстро принимают новый статус ученика, новые требования и режим дня.
Адаптация 3-4 месяца. Эти школьники могут не реагировать на замечания педагога, играть на уроках, могут обижаться, в усвоении учебной программы они испытывают трудности. Лишь к концу 2-ой четверти дети осваиваются и начинают вести себя адекватно в соответствии с требованиями школы и учителя.
Трудности в социально-психологической адаптации. У таких школьников может отмечаться крайне негативное поведение, нежелание слушаться, выполнять требования учителя. Они могут вступать в спор с педагогами, в драки со сверстниками. Усвоение учебного материала происходит с большим трудом. Как показывает практика, труднее всего адаптируются к новым условиям дети с СДВГ. Им необычайно сложно сидеть за партой и концентрировать свое внимание на учебном процессе. Они могут вскакивать во время урока, громко говорить, вставать и ходить по классу. Часто такие дети не чувствуют дистанцию между собой и учителем.
Они могут вступать в спор с педагогами, в драки со сверстниками. Усвоение учебного материала происходит с большим трудом. Как показывает практика, труднее всего адаптируются к новым условиям дети с СДВГ. Им необычайно сложно сидеть за партой и концентрировать свое внимание на учебном процессе. Они могут вскакивать во время урока, громко говорить, вставать и ходить по классу. Часто такие дети не чувствуют дистанцию между собой и учителем.
Физиологическая адаптация первоклассников занимает 1,5-2 месяца.В первом классе на плечи ребенка сваливается непривычная для него нагрузка. Какую бы работу ни выполнял школьник: умственную(выполняя задания), статическую(сидя в неподвижном положении за партой), психологическую (общение в коллективе) — на все это детский организм реагирует напряженной работой. Поэтому взрослым нужно следить за тем, чтобы ребенок регулярно менял положение тела, двигался (физкультминутки). Чтобы была частая смена деятельности, ведь время особого внимания 7 летнего ребёнка -10 -12 минут.
Признаки успешной адаптации:
- Удовлетворенность процессом обучения. Ребенку нравится в школе, он уверен в себе и не испытывает чувство страха.
- Ребенок справляется с учебной программой.
- Ребенок способен самостоятельно выполнять учебные задания, а при возникновении трудностей прибегать к помощи взрослых только после попыток справиться с задачей самому.
- Удовлетворенность ребенка межличностными отношениями с одноклассниками и учителями.
«Признаки адаптации». Экономист об уровне инфляции в РФ
https://radiosputnik.ria.ru/20220901/inflyatsiya-1813686351.html
«Признаки адаптации». Экономист об уровне инфляции в РФ
«Признаки адаптации». Экономист об уровне инфляции в РФ
«Признаки адаптации». Экономист об уровне инфляции в РФ
Радио Sputnik, 01. 09.2022
09.2022
2022-09-01T12:24
2022-09-01T12:24
2022-09-01T12:32
сказано в эфире
экономика
федеральная служба государственной статистики (росстат)
министерство экономического развития рф (минэкономразвития россии)
цены на нефть
россия
инфляция
подкасты – радио sputnik
/html/head/meta[@name=’og:title’]/@content
/html/head/meta[@name=’og:description’]/@content
https://cdnn21.img.ria.ru/images/152588/67/1525886730_0:91:6000:3466_1920x0_80_0_0_4464e1fea9a5a1e461576c68284dc0fe.jpg
Энергетический кризис в Европе, инфляция и рекомендации МВФ
— Франция прогнозирует улучшение ситуация с ростом цен и тарифов только в 2023 году.
— В МВФ посоветовали центробанкам усилить борьбу с ростом инфляции.
— Италия по примеру соседей по ЕС готовится к ограничению потребления газа и света.
— Немцы всё меньше доверяют экономической политике своих властей: показатели бьют антирекорды.
Эти и другие важные международные экономические события недели обсуждаем в эфире радио Sputnik.
audio/mpeg
Недельная дефляция в России в период с 23 по 29 августа ускорилась до 0,16%, подсчитали в Росстате. В интервью радио Sputnik кандидат экономических наук Михаил Беляев прокомментировал ситуацию.Росстат зафиксировал в стране падение потребительских цен, за неделю с 23 по 29 августа дефляция составила 0,16% после 0,15% за неделю с 16 по 22 августа. Об этом говорится в отчете Минэкономразвития. В годовом выражении инфляция замедлилась до 14,31%, отметили в ведомстве.В интервью радио Sputnik кандидат экономических наук, финансовый аналитик Михаил Беляев выразил мнение, что это не дефляция. По мере восстановления экономики уровень инфляции может опуститься ниже 10% в годовом выражении, прогнозирует эксперт. Коротко и по делу. Только отборные цитаты в нашем Телеграм-канале.
https://radiosputnik.ria.ru/20220831/ptitsa-1813366728.html
https://radiosputnik.ria.ru/20220901/top-menedzhery-1813544654.html
россия
Радио Sputnik
1
5
4. 7
7
96
7 495 645-6601
ФГУП МИА «Россия сегодня»
https://xn--c1acbl2abdlkab1og.xn--p1ai/awards/
2022
Радио Sputnik
1
5
4.7
96
7 495 645-6601
ФГУП МИА «Россия сегодня»
https://xn--c1acbl2abdlkab1og.xn--p1ai/awards/
Новости
ru-RU
https://radiosputnik.ria.ru/docs/about/copyright.html
https://xn--c1acbl2abdlkab1og.xn--p1ai/
Радио Sputnik
1
5
4.7
96
7 495 645-6601
ФГУП МИА «Россия сегодня»
https://xn--c1acbl2abdlkab1og.xn--p1ai/awards/
1920
1080
true
1920
1440
true
https://cdnn21.img.ria.ru/images/152588/67/1525886730_630:0:5370:3555_1920x0_80_0_0_0a5153cf8fbf811bfbc46c0c36f252e9.jpg
1920
1920
true
Радио Sputnik
1
5
4. 7
7
96
7 495 645-6601
ФГУП МИА «Россия сегодня»
https://xn--c1acbl2abdlkab1og.xn--p1ai/awards/
Радио Sputnik
1
5
4.7
96
7 495 645-6601
ФГУП МИА «Россия сегодня»
https://xn--c1acbl2abdlkab1og.xn--p1ai/awards/
экономика, федеральная служба государственной статистики (росстат), министерство экономического развития рф (минэкономразвития россии), цены на нефть, россия, инфляция, подкасты – радио sputnik
Сказано в эфире, Экономика, Федеральная служба государственной статистики (Росстат), Министерство экономического развития РФ (Минэкономразвития России), Цены на нефть, Россия, Инфляция, Подкасты – Радио Sputnik
Недельная дефляция в России в период с 23 по 29 августа ускорилась до 0,16%, подсчитали в Росстате. В интервью радио Sputnik кандидат экономических наук Михаил Беляев прокомментировал ситуацию.
31 августа, 11:27Сказано в эфире
«Насыщен и стабилен». Росптицесоюз о ситуации на рынке
Росптицесоюз о ситуации на рынке
Росстат зафиксировал в стране падение потребительских цен, за неделю с 23 по 29 августа дефляция составила 0,16% после 0,15% за неделю с 16 по 22 августа. Об этом говорится в отчете Минэкономразвития. В годовом выражении инфляция замедлилась до 14,31%, отметили в ведомстве.
В интервью радио Sputnik кандидат экономических наук, финансовый аналитик Михаил Беляев выразил мнение, что это не дефляция.
«Это не дефляция, а снижение темпов роста инфляции. Конечно, это хорошо. Инфляция связана с тем, какое состояние экономики в стране в данный момент фиксируется. Сейчас экономика находится в стабильном положении, и даже есть определенные признаки ее адаптации к новым условиям и признаки того, что она в ближайшее время может выйти на умеренные темпы роста, на положительную траекторию. Все это естественно отражается на макроэкономических показателях, в том числе, а может быть и в первую очередь на инфляции. Когда в экономике происходит нормализация процессов, нормализуются и тенденции по росту цен», – сказал он.
По мере восстановления экономики уровень инфляции может опуститься ниже 10% в годовом выражении, прогнозирует эксперт.
«Сейчас было зафиксировано где-то 14 процентов инфляции на годовом уровне, это еще очень высокий темп роста, но, по мере того, как будет налаживаться состояние экономики, она перейдет к ростовым характеристикам, думаю, инфляция будет приведена в норму, во всяком случае ниже 10 процентов опустится. Может, на какие-нибудь 5-6 процентов, которые сейчас уже инфляцией и не считаются. Если в экономике все будет в порядке, экономика будет развиваться, ни о каком росте инфляции говорить не придется», – считает Михаил Беляев.
1 сентября, 11:01В России
Российские топ-менеджеры ответят на вопросы о бизнесе
Коротко и по делу. Только отборные цитаты в нашем Телеграм-канале.
Психологическая адаптация человека
Адаптация личности – это психологическое приспособление к изменениям окружающей среды. Основная составляющая психологической адаптации заключается в приспособлении человека к требованиям социума и собственным желаниям. Психологическая адаптация подразумевает усвоение традиций социальной группы и принятых здесь ценностей. Она встречается повсеместно. Нам необходимо адаптироваться к ситуации в детском саду, в школе, на работе, в незнакомой компании.
Основная составляющая психологической адаптации заключается в приспособлении человека к требованиям социума и собственным желаниям. Психологическая адаптация подразумевает усвоение традиций социальной группы и принятых здесь ценностей. Она встречается повсеместно. Нам необходимо адаптироваться к ситуации в детском саду, в школе, на работе, в незнакомой компании.
Проявление и способы психологической адаптации
Психологическая адаптация – процесс, происходящий с раннего детства и до конца существования. Приспособление к окружающим условиям существования в детском возрасте чрезвычайно важно. Именно от психологической и социальной адаптации человека в детстве зависит его дальнейшая жизнь в социуме.
Все мы наслышаны об историях детей-Маугли. Человек, который в раннем возрасте адаптировался к диким условиям и жизни среди животных, не сможет вести активную социальную жизнь в обществе людей. Необходимость психологической адаптации к жизни среди себе подобных вызывает у Маугли шок. В редких случаях людям, выросшим без человеческого общества, удается приспособится к социуму. Чаще всего они возвращаются к прежней жизни, где психологическая адаптация – пройденный этап.
В редких случаях людям, выросшим без человеческого общества, удается приспособится к социуму. Чаще всего они возвращаются к прежней жизни, где психологическая адаптация – пройденный этап.
Главное проявление психологической адаптации — общение и другие виды взаимодействия. Активное взаимодействие с обществом и привыкание к нормам и устоям делают возможными процесс обучения и работы, построение отношений с другими членами общества, изменение поведения человека для соответствия ожиданиям окружающих.
Способы психологической адаптации:
Метод проб и ошибок. Встречаясь с жизненным препятствием на пути, человек преодолевает его, опираясь на собственный опыт. Не всегда испробованные способы преодоления решают возникшую проблему. Со временем человек отбрасывает в сторону методы, которые не привели к нужному результату, и находит новые пути решения.
Формирование реакции. Так называемая своеобразная «дрессировка». Правильная реакция на изменения общества подкрепляется вознаграждением на психологическом или физическом уровне. Этот способ адаптации не осознано используют родители в отношении детей. В тот момент, когда ребенок произносит членораздельные звуки, матери кажется, что он зовет именно ее. Она встречает начинание малыша с восторгом, что стимулирует ребенка к дальнейшему развитию.
Этот способ адаптации не осознано используют родители в отношении детей. В тот момент, когда ребенок произносит членораздельные звуки, матери кажется, что он зовет именно ее. Она встречает начинание малыша с восторгом, что стимулирует ребенка к дальнейшему развитию.
Наблюдение. Попадая в незнакомую среду, человек наблюдает за поведением окружающих. Он подражает людям, которые уже адаптировались к условиям взаимодействия в этом обществе, не задумываясь над тем, зачем он это делает. Со временем человек полностью принимает линию поведения, включая осознание целей и последствий.
Латентная адаптация. Человек постоянно получает сигналы от окружающего мира. Часть из них воспринимается, другая понимается не так четко, а третья не осознается вовсе. В подсознании сохраняются определенные методы взаимодействия с обществом, которые используются по мере необходимости и возникновения ситуаций.
Инсайт. Это реакция мозга на ситуацию, когда разбросанные в памяти варианты поведения и способы решения проблемы объединяются, и человек получает сигнал о том, какая реакция на окружающий мир станет единственно верной. Решение спонтанно и уникально, поэтому инсайт похож на творческий процесс.
Решение спонтанно и уникально, поэтому инсайт похож на творческий процесс.
Рассуждения. Обсуждая проблему или незнакомую ситуацию, человек ищет решение или способ адаптации к окружению. Выводы, полученные в результате рассуждений, в дальнейшем используются при аналогичных ситуациях.
Адаптационные расстройства личности
Не всегда процесс психологической адаптации человека проходит гладко. Если непривычная ситуация затягивается, то возможно появление адаптационных расстройств. Это может случится при затяжном семейном конфликте, при расставании с близкими людьми, при потери прежнего положения в социуме или при тяжелом заболевании.
Опасные звоночки проявляются постепенно. Развитие адаптационного расстройства занимает около месяца и в дальнейшем может либо исчезнуть, либо перерасти в психическое заболевание, требующее вмешательства врача.
Варианты адаптационного расстройства человека:
Кратковременная депрессия, которая заключается в резком понижении настроения, апатичности и безразличия, медлительности мышления, ощущении бессилия и неуверенности в себе;
Пролонгированная депрессия, которая подразумевает продолжительность симптомов кратковременной до двух лет. Проявляется, когда конфликтная жизненная ситуация затягивается, а человек не может с ней справится и адаптироваться к изменениям собственной жизни;
Проявляется, когда конфликтная жизненная ситуация затягивается, а человек не может с ней справится и адаптироваться к изменениям собственной жизни;
Тревожность, проявляющаяся в форме беспокойства, как двигательного, так и вегетативного;
Нарушение эмоционального фона, когда наряду с депрессией и тревожностью, проявляется агрессия, раздражительность, подверженность состоянию гнева и злости;
Нарушения поведения, проявляющееся в поступках, которые намеренно нарушают этические и моральные нормы, принятые в обществе. Такие отклонения характерны для подросткового возраста, когда необходимость психологической адаптации вызывает в человеке отрицание и подталкивает на бессмысленные действия.
Расстройства, связанные с психологической адаптацией человека, распространены в обществе. Однако подобный диагноз ставится врачом только в случае нарушения функциональности человека, как полноценного члена общества. Чаще всего адаптационными расстройствами личности страдают женщины. Среди людей, которые не смогли психологически адаптироваться к происходящему, преобладают те, которые считают себя несчастными. Это одинокие люди, не имеющие семьи или разведенные, социальное и материальное положение человека с расстройством, как правило, оставляет желать лучшего.
Среди людей, которые не смогли психологически адаптироваться к происходящему, преобладают те, которые считают себя несчастными. Это одинокие люди, не имеющие семьи или разведенные, социальное и материальное положение человека с расстройством, как правило, оставляет желать лучшего.
Адаптационные расстройства успешно поддаются лечению и в большинстве случаев имеют благоприятный исход, если вовремя обратится к специалисту и принять меры. Психологическая адаптация – обязательный процесс для гармоничного существования в обществе и общения с его членами.
Адаптация и выживание | Национальное географическое общество
Любое количество характеристик может различаться у особей данного вида — некоторые могут быть крупнее, мохнатее, лучше бороться с инфекциями или иметь меньшие уши. Эти характеристики в значительной степени определяются их генами, которые передаются от родителей и впоследствии передаются их собственному потомству. Некоторые из этих характеристик или черт обеспечивают конкурентные преимущества, такие как скорость, сила или привлекательность. Если эти черты особенно полезны, люди с этими чертами произведут больше потомства, чем те, у кого их нет. С течением поколений количество особей с этим выгодным признаком или адаптацией будет увеличиваться, пока он не станет общим признаком вида.
Если эти черты особенно полезны, люди с этими чертами произведут больше потомства, чем те, у кого их нет. С течением поколений количество особей с этим выгодным признаком или адаптацией будет увеличиваться, пока он не станет общим признаком вида.
Структурные и поведенческие адаптации
Адаптация может быть структурной, то есть физической частью организма. Адаптация также может быть поведенческой, влияющей на то, как организм реагирует на окружающую среду.
Пример структурной адаптации – то, как некоторые растения приспособились к жизни в сухих жарких пустынях. Растения, называемые суккулентами, приспособились к этому климату, накапливая воду в своих коротких толстых стеблях и листьях.
Сезонная миграция — пример поведенческой адаптации. Серые киты ( Eschrichtius robustus ) ежегодно мигрируют на тысячи километров, переплывая летом из холодного Северного Ледовитого океана в теплые воды у побережья Мексики на зиму. Детеныши серых китов рождаются в теплых южных водах, а затем перемещаются группами, называемыми стаями, в богатые питательными веществами воды Арктики.
Адаптации, развивающиеся в ответ на одну проблему, иногда помогают или используются для решения другой. Перья, вероятно, были первым приспособлением для осязания или регулирования температуры. Позже перья стали длиннее и жестче, позволяя планировать, а затем и летать. Такие признаки называются экзаптациями.
Некоторые черты, с другой стороны, теряют свою функцию, когда другие приспособления становятся более важными или когда меняется окружающая среда. Свидетельства этих черт остаются в рудиментарной форме – уменьшены или бесфункциональны. У китов и дельфинов есть рудиментарные кости ног, остатки приспособления (ноги), которые их предки использовали для ходьбы.
Среда обитания
Адаптации часто развиваются в ответ на изменение среды обитания организмов.
Знаменитым примером приспособления животного к изменению окружающей среды является английская бабочка перцовая ( Biston betularia ). До 19 века самый распространенный вид этой бабочки был кремового цвета с более темными пятнами. Несколько перченых мотыльков были серыми или черными.
Несколько перченых мотыльков были серыми или черными.
Промышленная революция изменила окружающую среду, изменился и внешний вид перцовой моли. Мотыльки более темного цвета, встречавшиеся редко, начали размножаться в городской атмосфере. Их сажевый цвет сливался с деревьями, запачканными промышленными загрязнениями. Птицы также не могли видеть темных бабочек, поэтому вместо них они ели кремовых бабочек. Мотыльки кремового цвета начали возвращаться после того, как Великобритания приняла законы, ограничивающие загрязнение воздуха.
Видообразование
Иногда развивается адаптация или набор адаптаций, которые разделяют один вид на два. Этот процесс известен как видообразование.
Сумчатые в Океании являются примером адаптивной радиации, типа видообразования, при котором виды развиваются, чтобы заполнить множество пустых экологических ниш. Сумчатые, млекопитающие, вынашивающие своих развивающихся детенышей в сумках после непродолжительной беременности, прибыли в Океанию до того, как земля отделилась от Азии. Плацентарные млекопитающие, животные, вынашивающие своих детенышей в утробе матери, стали доминировать на всех остальных континентах, но не в Океании. Коалы ( Phascolarctos cinereus ), например, приспособился питаться эвкалиптовыми деревьями, произрастающими в Австралии. Вымерший тасманийский тигр ( Thylacinus cynocephalus ) был плотоядным сумчатым и приспособился к нише, заполненной большими кошками, такими как тигры, на других континентах.
Плацентарные млекопитающие, животные, вынашивающие своих детенышей в утробе матери, стали доминировать на всех остальных континентах, но не в Океании. Коалы ( Phascolarctos cinereus ), например, приспособился питаться эвкалиптовыми деревьями, произрастающими в Австралии. Вымерший тасманийский тигр ( Thylacinus cynocephalus ) был плотоядным сумчатым и приспособился к нише, заполненной большими кошками, такими как тигры, на других континентах.
Цихлиды, обитающие во многих озерах Африки, демонстрируют другой тип видообразования, симпатрическое видообразование. Симпатрическое видообразование противоположно физической изоляции. Это происходит, когда виды разделяют одну и ту же среду обитания. Адаптация позволила сотням разновидностей цихлид жить в озере Малави. Каждый вид цихлид имеет уникальную специализированную диету: один вид цихлид может питаться только насекомыми, другой может питаться только водорослями, третий может питаться только другой рыбой.
Коадаптация
Организмы иногда адаптируются с другими организмами и к ним. Это называется коадаптацией. Некоторые цветы производят нектар, который нравится колибри. Колибри, в свою очередь, приспособили длинные тонкие клювы, чтобы извлекать нектар из определенных цветов. Когда колибри идет на кормежку, она непреднамеренно собирает пыльцу с пыльников цветов, которая оседает на рыльце следующих цветов, которые она посещает. В этих отношениях колибри получает пищу, а пыльца растений распространяется. Коадаптация полезна для обоих организмов.
Это называется коадаптацией. Некоторые цветы производят нектар, который нравится колибри. Колибри, в свою очередь, приспособили длинные тонкие клювы, чтобы извлекать нектар из определенных цветов. Когда колибри идет на кормежку, она непреднамеренно собирает пыльцу с пыльников цветов, которая оседает на рыльце следующих цветов, которые она посещает. В этих отношениях колибри получает пищу, а пыльца растений распространяется. Коадаптация полезна для обоих организмов.
Мимикрия — еще один вид коадаптации. В мимикрии один организм приспособился быть похожим на другой. Безобидная королевская змея (иногда называемая молочной змеей) адаптировала цветовой узор, напоминающий смертоносную коралловую змею. Эта мимикрия удерживает хищников от королевской змеи.
Мимический осьминог ( Thaumoctopus mimicus ) имеет поведенческие и структурные адаптации. Этот вид осьминогов может копировать внешний вид и движения других животных, таких как морские змеи, камбалы, медузы и креветки.
Коадаптация также может ограничивать способность организма приспосабливаться к новым изменениям в среде обитания. Это может привести к совместному вымиранию. В южной Англии большая голубая бабочка приспособилась есть красных муравьев. Когда развитие человека сократило среду обитания красных муравьев, локальное исчезновение красных муравьев привело к локальному исчезновению большой синей бабочки.
Статьи и профили
Новости National Geographic: три высокогорных народа, три приспособления к разреженной атмосфере Новости National Geographic: открытие ископаемых четвероногих — от плавников до конечностей
Интерактивы
День Земли, Канада: Адаптация животных
Признаки адаптации к местным условиям рН в мозаике окружающей среды в современной экосистеме Калифорнии
Обзор
. 2013 ноябрь; 53(5):857-70.
doi: 10.1093/icb/ict094.
Epub 2013 26 августа.
М Х Песпени 1 , Ф. Чан, Б. А. Менге, С. Р. Палумби,
Принадлежности
принадлежность
- 1 *Кафедра биологии, Университет Индианы, Блумингтон, IN 47405, США; Кафедра зоологии Орегонского государственного университета, Корваллис, штат Орегон, 97331, США; Факультет биологии Стэнфордского университета, морская станция Хопкинс, Пасифик Гроув, Калифорния
, США.
- PMID: 23980118
- DOI: 10.1093/icb/ict094
Обзор
M H Pespeni et al. Интегр Комп Биол. 2013 ноябрь
. 2013 ноябрь; 53(5):857-70.
2013 ноябрь; 53(5):857-70.
doi: 10.1093/icb/ict094. Epub 2013 26 августа.
Авторы
М Х Песпени 1 , Ф. Чан, Б. А. Менге, С. Р. Палумби
принадлежность
- 1 *Кафедра биологии, Университет Индианы, Блумингтон, IN 47405, США; Кафедра зоологии Орегонского государственного университета, Корваллис, штат Орегон, 97331, США; Факультет биологии Стэнфордского университета, морская станция Хопкинс, Пасифик Гроув, Калифорния
, США.
- PMID: 23980118
- DOI: 10.1093/icb/ict094
Абстрактный
Мало что известно о возможности быстрой эволюции естественных популяций в ответ на высокие темпы современных климатических изменений.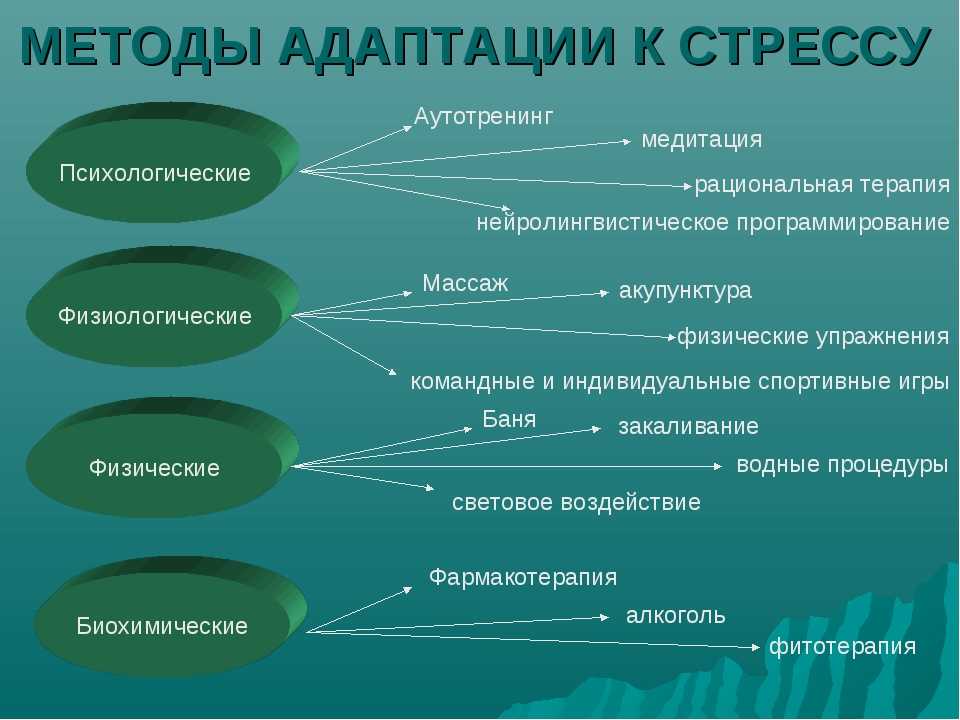 Особый интерес представляют организмы, развившиеся в средах с высокой изменчивостью в пространстве и времени, поскольку они могут содержать генетические вариации, которые могут способствовать эволюционной реакции на меняющиеся условия. Здесь мы рассмотрим, что известно о генетической способности к адаптации пурпурного морского ежа, Strongylocentrotus purpuratus, вида, который эволюционировал в апвеллинговой экосистеме северо-восточной части Тихого океана. Мы также представляем новые результаты тестирования адаптации к местным условиям рН в шести популяциях от Орегона до южной Калифорнии. Интегрируем данные по 19,493 генетических полиморфизма с данными о местных условиях рН. Мы обнаружили корреляции между частотой аллелей и средним ранговым временем, проведенным при pH <7,8, в 318 однонуклеотидных полиморфизмах в 275 генах. Два гена, наиболее коррелирующие с локальным pH, представляют собой белок, связанный с цитоскелетом и протонным насосом, с функциональной ролью в поддержании объема клетки и внутренней регуляции pH соответственно.
Особый интерес представляют организмы, развившиеся в средах с высокой изменчивостью в пространстве и времени, поскольку они могут содержать генетические вариации, которые могут способствовать эволюционной реакции на меняющиеся условия. Здесь мы рассмотрим, что известно о генетической способности к адаптации пурпурного морского ежа, Strongylocentrotus purpuratus, вида, который эволюционировал в апвеллинговой экосистеме северо-восточной части Тихого океана. Мы также представляем новые результаты тестирования адаптации к местным условиям рН в шести популяциях от Орегона до южной Калифорнии. Интегрируем данные по 19,493 генетических полиморфизма с данными о местных условиях рН. Мы обнаружили корреляции между частотой аллелей и средним ранговым временем, проведенным при pH <7,8, в 318 однонуклеотидных полиморфизмах в 275 генах. Два гена, наиболее коррелирующие с локальным pH, представляют собой белок, связанный с цитоскелетом и протонным насосом, с функциональной ролью в поддержании объема клетки и внутренней регуляции pH соответственно. Во всех протестированных локусах высокая корреляция с локальным рН была сосредоточена в генах, связанных с транспортом ионов, биоминерализацией, метаболизмом липидов и межклеточной адгезией, функциональными путями, важными для поддержания гомеостаза при низком рН. Мы определили набор из семи генов в качестве главных кандидатов на быструю эволюционную реакцию на закисление океана. В этих генах предполагаемый адаптированный к низкому pH аллель, основанный на частотах аллелей в естественных популяциях, быстро увеличивается по частоте у личинок пурпурных морских ежей, выращенных при низком pH. Мы также обнаружили, что популяции из местностей с высоким pH показывают большее изменение частоты аллелей в сторону предполагаемых адаптированных к низкому pH аллелей при экспериментальном подкислении по сравнению с популяциями с низким pH, что свидетельствует о том, что как естественный, так и искусственный отбор отдают предпочтение одним и тем же аллелям для реакции на низкий рН. Эти результаты показывают, что пурпурные морские ежи могут быть адаптированы к местному pH, и предполагают, что этот вид может обладать генетической способностью к быстрой эволюции в ответ на закисление.
Во всех протестированных локусах высокая корреляция с локальным рН была сосредоточена в генах, связанных с транспортом ионов, биоминерализацией, метаболизмом липидов и межклеточной адгезией, функциональными путями, важными для поддержания гомеостаза при низком рН. Мы определили набор из семи генов в качестве главных кандидатов на быструю эволюционную реакцию на закисление океана. В этих генах предполагаемый адаптированный к низкому pH аллель, основанный на частотах аллелей в естественных популяциях, быстро увеличивается по частоте у личинок пурпурных морских ежей, выращенных при низком pH. Мы также обнаружили, что популяции из местностей с высоким pH показывают большее изменение частоты аллелей в сторону предполагаемых адаптированных к низкому pH аллелей при экспериментальном подкислении по сравнению с популяциями с низким pH, что свидетельствует о том, что как естественный, так и искусственный отбор отдают предпочтение одним и тем же аллелям для реакции на низкий рН. Эти результаты показывают, что пурпурные морские ежи могут быть адаптированы к местному pH, и предполагают, что этот вид может обладать генетической способностью к быстрой эволюции в ответ на закисление. Эта адаптивная способность, вероятно, возникает из-за постоянной генетической изменчивости, поддерживаемой в природе путем уравновешивания отбора в пространственной и временной мозаике окружающей среды, которая характеризует Калифорнийскую текущую экосистему.
Эта адаптивная способность, вероятно, возникает из-за постоянной генетической изменчивости, поддерживаемой в природе путем уравновешивания отбора в пространственной и временной мозаике окружающей среды, которая характеризует Калифорнийскую текущую экосистему.
Похожие статьи
Транскриптомные реакции на подкисление морской воды среди популяций морских ежей, населяющих естественную мозаику рН.
Эванс Т.Г., Песпени М.Х., Хофманн Г.Е., Палумби С.Р., Сэнфорд Э. Эванс Т.Г. и соавт. Мол Экол. 2017 апр; 26 (8): 2257-2275. doi: 10.1111/mec.14038. Epub 2017 8 марта. Мол Экол. 2017. PMID: 28141889
Эволюционные изменения во время экспериментального закисления океана.
Песпени М.Х., Сэнфорд Э., Гейлорд Б.
 , Хилл Т.М., Хосфельт Д.Д., Ярис Х.К., ЛаВинь М., Ленц Э.А., Рассел А.Д., Янг М.К., Палумби С.Р.
Песпени М.Х. и соавт.
Proc Natl Acad Sci USA. 2013 Apr 23;110(17):6937-42. doi: 10.1073/pnas.1220673110. Epub 2013 8 апр.
Proc Natl Acad Sci U S A. 2013.
PMID: 23569232
Бесплатная статья ЧВК.
, Хилл Т.М., Хосфельт Д.Д., Ярис Х.К., ЛаВинь М., Ленц Э.А., Рассел А.Д., Янг М.К., Палумби С.Р.
Песпени М.Х. и соавт.
Proc Natl Acad Sci USA. 2013 Apr 23;110(17):6937-42. doi: 10.1073/pnas.1220673110. Epub 2013 8 апр.
Proc Natl Acad Sci U S A. 2013.
PMID: 23569232
Бесплатная статья ЧВК.Естественная изменчивость и способность адаптироваться к закислению океана у краеугольного морского ежа Strongylocentrotus purpuratus.
Келли М.В., Падилья-Гаминьо Х.Л., Хофманн Г.Э. Келли М.В. и др. Глоб Чанг Биол. 2013 авг; 19 (8): 2536-46. doi: 10.1111/gcb.12251. Epub 2013 11 июня. Глоб Чанг Биол. 2013. PMID: 23661315
Исследования подкисления океана в «постгеномную» эпоху: дорожные карты пурпурного морского ежа Strongylocentrotus purpuratus.

Evans TG, Padilla-Gamiño JL, Kelly MW, Pespeni MH, Chan F, Menge BA, Gaylord B, Hill TM, Russell AD, Palumbi SR, Sanford E, Hofmann GE. Эванс Т.Г. и соавт. Comp Biochem Physiol A Mol Integr Physiol. 2015 июль; 185:33-42. doi: 10.1016/j.cbpa.2015.03.007. Epub 2015 13 марта. Comp Biochem Physiol A Mol Integr Physiol. 2015. PMID: 25773301 Обзор.
Географические градиенты в отборе могут выявить генетические ограничения для эволюционных реакций на закисление океана.
Гайтан-Эспития Д.Д., Маршалл Д., Дюпон С., Басигалупе Л.Д., Бодросси Л., Хобдай А.Дж. Gaitán-Espitia JD, et al. Биол Летт. 2017 Февраль;13(2):20160784. doi: 10.1098/rsbl.2016.0784. Биол Летт. 2017. PMID: 28148831 Бесплатная статья ЧВК. Обзор.
Посмотреть все похожие статьи
Цитируется
Индивидуальные экоэволюционные модели для понимания адаптации к изменяющимся морям.

Сюереб А., Ружмонт К., Тиффин П., Сюэ Х., Файфер-Рикси М. Сюреб А. и др. Proc Biol Sci. 2021 10 ноября; 288 (1962): 20212006. doi: 10.1098/rspb.2021.2006. Epub 2021 10 ноября. Proc Biol Sci. 2021. PMID: 34753353 Обзор.
Паттерны экспрессии генов у красных морских ежей (Mesocentrotus franciscanus), подвергшихся воздействию различных комбинаций температуры и pCO 2 на раннем этапе развития.
Вонг Дж. М., Хофманн Г. Э. Вонг Дж.М. и др. Геномика BMC. 2021 7 января; 22(1):32. doi: 10.1186/s12864-020-07327-x. Геномика BMC. 2021. PMID: 33413121 Бесплатная статья ЧВК.
Подкисление океана способствует широкому транскрипционному ответу морских многоклеточных животных: обзор литературы.

Страдер М.Э., Вонг Дж.М., Хофманн Г.Э. Страдер М.Е. и др. Фронт Зоол. 2020 17 фев; 17:7. дои: 10.1186/с12983-020-0350-9. Электронная коллекция 2020. Фронт Зоол. 2020. PMID: 320
Бесплатная статья ЧВК. Обзор.Влияние изменения климата на кинематику побега и поведение рыб: последствия для будущих взаимодействий хищник-жертва.
Доменичи П., Аллан Б.Дж.М., Лефрансуа С., Маккормик М.И. Доменичи П. и др. Консерв Физиол. 2019 ноябрь 7;7(1):coz078. doi: 10.1093/conphys/coz078. электронная коллекция 2019. Консерв Физиол. 2019. PMID: 31723432 Бесплатная статья ЧВК.
Влияние пониженного pH на физиологию и целостность скорлупы молоди Haliotis iris (pāua) из Новой Зеландии.
Каммингс В.
 Дж., Смит А.М., Марриотт П.М., Пиблз Б.А., Холлидей, Нью-Джерси.
Каммингс В.Дж. и др.
Пир Дж. 2019 25 сентября; 7: e7670. doi: 10.7717/peerj.7670. Электронная коллекция 2019.
Пир Дж. 2019.
PMID: 31579589Бесплатная статья ЧВК.
Дж., Смит А.М., Марриотт П.М., Пиблз Б.А., Холлидей, Нью-Джерси.
Каммингс В.Дж. и др.
Пир Дж. 2019 25 сентября; 7: e7670. doi: 10.7717/peerj.7670. Электронная коллекция 2019.
Пир Дж. 2019.
PMID: 31579589Бесплатная статья ЧВК.
Просмотреть все статьи «Цитируется по»
Типы публикаций
термины MeSH
Смягчение последствий и адаптация | Решения – изменение климата: жизненно важные признаки планеты
НАСА является мировым лидером в области изучения климата и наук о Земле. Хотя его роль не заключается в установлении климатической политики или предписании конкретных ответных мер или решений в связи с изменением климата, в его компетенцию входит предоставление надежных научных данных, необходимых для понимания изменения климата. Затем НАСА предоставляет эту информацию мировому сообществу – общественности, политикам и лицам, принимающим решения, а также научным и планирующим агентствам по всему миру.
Затем НАСА предоставляет эту информацию мировому сообществу – общественности, политикам и лицам, принимающим решения, а также научным и планирующим агентствам по всему миру.
Изменение климата — одна из самых сложных проблем, стоящих перед нами сегодня. Она включает в себя множество аспектов — науку, экономику, общество, политику и морально-этические вопросы — и представляет собой глобальную проблему, ощущаемую в локальных масштабах, которая будет существовать тысячи лет. Углекислый газ, улавливающий тепло парниковый газ, который является основной причиной недавнего глобального потепления, задерживается в атмосфере на многие тысячи лет, и планете (особенно океану) требуется время, чтобы отреагировать на потепление. Таким образом, даже если сегодня мы прекратим выбросы всех парниковых газов, глобальное потепление и изменение климата продолжат влиять на будущие поколения. Таким образом, человечество «привержено» определенному уровню изменения климата.
Насколько изменится климат? Это будет определяться тем, как будут продолжаться наши выбросы и как именно наш климат реагирует на эти выбросы. Несмотря на растущую осведомленность об изменении климата, наши выбросы парниковых газов продолжают неуклонно расти. В 2013 году дневной уровень углекислого газа в атмосфере впервые в истории человечества превысил 400 частей на миллион. В последний раз уровни были такими высокими примерно от трех до пяти миллионов лет назад, в эпоху плиоцена.
Поскольку мы уже привержены определенному уровню изменения климата, реагирование на изменение климата включает двусторонний подход:
- Сокращение выбросов и стабилизация уровней удерживающих тепло парниковых газов в атмосфере («смягчение последствий») ;
- Адаптация к изменению климата уже в разработке («адаптация») .
Смягчение последствий — уменьшение изменения климата — включает уменьшение потока удерживающих тепло парниковых газов в атмосферу либо за счет сокращения источников этих газов (например, сжигание ископаемого топлива для производства электроэнергии, тепла или транспорта), либо за счет увеличения «поглотители», которые аккумулируют и хранят эти газы (такие как океаны, леса и почва). Цель смягчения последствий состоит в том, чтобы избежать значительного вмешательства человека в климат Земли, «стабилизировать уровни парниковых газов в сроки, достаточные для того, чтобы позволить экосистемам естественным образом адаптироваться к изменению климата, гарантировать, что производство продуктов питания не будет поставлено под угрозу, и позволить экономическому развитию продолжаться в устойчивым образом» (из отчета Межправительственной группы экспертов ООН по изменению климата за 2014 г. о смягчении последствий изменения климата, стр. 4).
Цель смягчения последствий состоит в том, чтобы избежать значительного вмешательства человека в климат Земли, «стабилизировать уровни парниковых газов в сроки, достаточные для того, чтобы позволить экосистемам естественным образом адаптироваться к изменению климата, гарантировать, что производство продуктов питания не будет поставлено под угрозу, и позволить экономическому развитию продолжаться в устойчивым образом» (из отчета Межправительственной группы экспертов ООН по изменению климата за 2014 г. о смягчении последствий изменения климата, стр. 4).
Адаптация – приспособление к жизни в изменяющемся климате – включает приспособление к фактическому или ожидаемому будущему климату. Цель состоит в том, чтобы снизить риски, связанные с вредными последствиями изменения климата (такими как повышение уровня моря, более интенсивные экстремальные погодные явления или отсутствие продовольственной безопасности). Это также включает максимальное использование любых потенциальных выгодных возможностей, связанных с изменением климата (например, более продолжительный вегетационный период или повышение урожайности в некоторых регионах).
На протяжении всей истории люди и общества приспосабливались и справлялись с изменениями климата и экстремальными явлениями с разной степенью успеха. Изменение климата (в частности, засуха) было, по крайней мере, частично ответственным за подъем и падение цивилизаций. Климат Земли был относительно стабильным в течение последних 10 000 лет, и эта стабильность способствовала развитию нашей современной цивилизации и сельского хозяйства. Наша современная жизнь приспособлена к этому стабильному климату, а не к гораздо более теплому климату следующей тысячи с лишним лет. Поскольку наш климат меняется, нам нужно будет адаптироваться. Чем быстрее меняется климат, тем сложнее будет.
Хотя изменение климата является глобальной проблемой, оно ощущается в локальном масштабе. Таким образом, местные органы власти находятся на переднем крае адаптации. Города и местные сообщества по всему миру сосредоточили свое внимание на решении собственных климатических проблем. Они работают над созданием защиты от наводнений, планированием периодов сильной жары и более высоких температур, укладкой тротуаров с улучшенным дренажом для борьбы с наводнениями и ливневыми водами, а также над улучшением хранения и использования воды.
Согласно отчету Межправительственной группы экспертов ООН по изменению климата ООН о воздействии изменения климата, адаптации и уязвимости за 2014 год (стр. 8), правительства разных уровней также лучше справляются с адаптацией. Изменение климата включается в планы развития: как справляться со все более экстремальными бедствиями, которые мы наблюдаем, как защищать береговые линии и бороться с повышением уровня моря, как лучше всего управлять землей и лесами, как бороться с засухой и планировать ее, как для разработки новых сортов сельскохозяйственных культур и как защитить энергию и общественную инфраструктуру.
Участие НАСА НАСА с его взглядом на Землю и обширными знаниями о климате Земли является одним из мировых экспертов в области климатологии. Роль НАСА заключается в предоставлении надежных научных данных, необходимых для понимания изменения климата. Например, данные эксперимента агентства по восстановлению гравитации и климата (GRACE), его последующей миссии (GRACE-FO), спутников для наблюдения за льдом, облаками и высотой суши (ICESat) и миссий ICESat-2 показали быстрые изменения в огромные ледяные щиты Земли. Серия миссий Sentinel-6 Майкла Фрейлиха и Джейсона зафиксировала повышение глобального уровня моря с 19 века.92.
Серия миссий Sentinel-6 Майкла Фрейлиха и Джейсона зафиксировала повышение глобального уровня моря с 19 века.92.
НАСА предоставляет подробные климатические данные мировому сообществу – общественности, политикам и лицам, принимающим решения, а также научным и планирующим агентствам по всему миру. В задачи НАСА не входит определение климатической политики или рекомендации решений по изменению климата. НАСА является одним из 13 правительственных агентств США, входящих в Программу исследований глобальных изменений США, которая имеет юридический мандат помогать нации и миру понимать, оценивать, прогнозировать и реагировать на глобальные изменения. Эти агентства-партнеры США включают Министерство сельского хозяйства, Агентство по охране окружающей среды и Министерство энергетики, каждое из которых играет свою роль в зависимости от своей области знаний.
Хотя основное внимание НАСА сосредоточено не на исследованиях и разработках в области энергетических технологий, в агентстве и с участием различных партнеров и сотрудников ведется работа по поиску других источников энергии для удовлетворения наших потребностей.
Для получения дополнительной информации о работе НАСА по смягчению последствий и адаптации посетите следующие страницы:
- Наука о Земле в действии
- Устойчивое развитие и государственные ресурсы
- . Электрический самолет НАСА .
- НАСА Аэронавтика
- Отделение НАСА (Программа передачи технологий)
- Реагирование на изменение климата включает два возможных подхода: снижение и стабилизация уровней удерживающих тепло парниковых газов в атмосфере («смягчение последствий») и адаптация к изменению климата, которое уже находится в разработке («адаптация») .
- Роль НАСА заключается в предоставлении климатических данных мировому сообществу, в том числе общественности, политикам и лицам, принимающим решения, а также научным и планирующим агентствам.
Видео: Годовой минимум морского льда в Арктике за 1979–2021 гг. с площадным графиком
Видео: глобальное потепление с 1880 по 2021 год
Видео: Ozone Watch 2018
Последние новости и возможности
Биологическая адаптируемость человека: обзор
| Инуиты (эскимосы), проживающие в полярной пустыне — пример человека способность выживать в экстремальные условия |
Человек
организм легко реагирует на изменяющиеся экологические стрессы разнообразными биологическими и
культурные пути. Мы можем акклиматизироваться к широкому диапазону температур и
влажность. При путешествии на большие высоты наши тела приспосабливаются так, что наши клетки
все еще получают достаточное количество кислорода. Мы также постоянно реагируем на физиологические
пути к внутренним и внешним стрессам, таким как бактериальные
и вирусные инфекции,
загрязнение воздуха и воды, питание
дисбаланс и перенаселенность.
Мы можем акклиматизироваться к широкому диапазону температур и
влажность. При путешествии на большие высоты наши тела приспосабливаются так, что наши клетки
все еще получают достаточное количество кислорода. Мы также постоянно реагируем на физиологические
пути к внутренним и внешним стрессам, таким как бактериальные
и вирусные инфекции,
загрязнение воздуха и воды, питание
дисбаланс и перенаселенность.
Эта способность быстро приспосабливаться к изменяющимся условиям окружающей среды позволяет нам, чтобы выжить в большинстве регионов мира. Мы успешно живем во влажных тропических леса, суровые пустыни, арктические пустоши и даже густонаселенные города с значительное количество загрязнений. Большинство других видов животных и растений ограничены одним или относительно немного сред из-за их более ограниченной приспособляемости.
Люди обычно реагируют на экологические стрессы четырьмя способами:
| биологический ответы | 1. генетическое изменение генетическое изменение | |||||||
|---|---|---|---|---|---|---|---|---|
| 2. корректировка развития | ответы без генетическое изменение | |||||||
| 3. Акклиматизация | ||||||||
| 4. культурные обычаи и технологии |
Первые три являются биологическими реакциями. Последние три происходят в течение нашей жизни. без дальнейших генетических изменений.
Генетика
Смена
Когда
экологический стресс постоянен и длится на протяжении многих поколений, успешная адаптация может
развиваться путем биологической эволюции. Те люди, которые наследуют признак, который
предлагает преимущество в реагировании на определенные стрессы
более
вероятно, проживут дольше и передадут больше своих генов следующему поколению.
Это эволюция через естественный отбор.
Например,
люди, чьи предки жили на территориях,
эндемичный
малярия
на протяжении тысячелетий часто наследуют некоторую степень иммунитета к этому серьезному заболеванию.
Высокая частота серповидно-клеточного признака
у народов Центральной Африки во многом является результатом косвенного отбора на это
признак малярии. Гетерозиготные носители серповидного гена обычно не имеют
серповидноклеточной анемии и достаточно устойчивы к малярийным
микроорганизмов, что они обладают избирательным преимуществом. Другой пример
генетическое решение стресса окружающей среды — наша способность производить пот
как помощь в охлаждении нашего тела в жарких условиях. Это не удивительно
что у нас есть эта возможность, потому что наша
непосредственные дочеловеческие предки были тропическими
животные.
Те люди, которые наследуют признак, который
предлагает преимущество в реагировании на определенные стрессы
более
вероятно, проживут дольше и передадут больше своих генов следующему поколению.
Это эволюция через естественный отбор.
Например,
люди, чьи предки жили на территориях,
эндемичный
малярия
на протяжении тысячелетий часто наследуют некоторую степень иммунитета к этому серьезному заболеванию.
Высокая частота серповидно-клеточного признака
у народов Центральной Африки во многом является результатом косвенного отбора на это
признак малярии. Гетерозиготные носители серповидного гена обычно не имеют
серповидноклеточной анемии и достаточно устойчивы к малярийным
микроорганизмов, что они обладают избирательным преимуществом. Другой пример
генетическое решение стресса окружающей среды — наша способность производить пот
как помощь в охлаждении нашего тела в жарких условиях. Это не удивительно
что у нас есть эта возможность, потому что наша
непосредственные дочеловеческие предки были тропическими
животные.
Генетические изменения в ответ на экологические стрессы обычно требуют много поколений, чтобы получили широкое распространение в популяции. К счастью, у нас есть и другие способы реагируя быстрее как личности в течение нашей собственной жизни. Слово настройки используется здесь для обозначения этих более коротких терминов физиологические изменения, которые не передаются по наследству. слово адаптации зарезервирован для наследственных генетических изменений, развившихся в популяции в течение длительного периода времени.
Развитие
Регулировка
Один из
более мощные виды приспособления к стрессам окружающей среды – это изменение роста
закономерности и развитие. Это происходит в детстве и обычно приводит к анатомическим
и/или физиологические
изменения, которые
в большинстве случаев необратимы во взрослом возрасте. Такие постоянные изменения называются
к как адаптация развития или адаптация развития .
| Рентген раннего
20 век Связанная ступня китаянки Рост остановился и |
Среди людей,
корректировки развития являются результатом как естественного давления окружающей среды, так и культурных факторов.
практики. Примером последнего был ныне незаконный обычай
в Китае туго обматывать или связывать ноги молодых
девушки с тканью, чтобы
препятствуют нормальному росту. В то время как это вызвало постоянные, калечащие деформации
кости стопы, это также привело к чрезвычайно крошечным стопам, которые считались очень
привлекательный. Родители искалечили своих дочерей из добрых побуждений.
Маленькие ноги сделают их более привлекательными партнерами по браку для богатых важных мужчин. и спаси их от тяжелой жизни.
и спаси их от тяжелой жизни.
| Конец 19 века британский актриса (Лилли Лэнгтри) кто типизировал идеал красота со своей осой- фигурная талия достигнуто с тугой корсет |
Легко осуждать старый китайский обычай связывания ног считается варварским. Однако стоит учитывая, что североамериканцы и европейцы намеренно измененные части тела своих детей и себя с неприятными процедуры тоже. В конце 19века, тугие корсеты, которые носят девочки, когда их тела еще росли, имели эффект деформации нижней реберные кости опасно приближаются к их легким. Некоторые богатые женщины даже имели нижние ребра удалены хирургическим путем, чтобы добиться стильной «осиной» формы Талия. Окружность 19 дюймов была идеальной.
Преднамеренная деформация частей тела — это не то, что случалось только в прошлом. Сегодня в Китае растет беспокойство среди многих высокомобильных средних компаний.
класс мужчин и женщин, что они слишком короткие. Тысячи искали
решение этой проблемы путем удлинения ног. Это
достигается долгим и болезненным процессом, включающим хирургическое разрушение
две кости голени в обеих ногах, а затем с помощью регулируемых металлических скоб, которые
закреплены стальными штифтами, имплантированными в кость чуть ниже колен и
над лодыжками, чтобы постепенно увеличить длину почти на 1/16 дюйма
(около 1 мм.) в сутки по мере срастания костей. Это увеличивает разрыв в
зоны перелома, тем самым стимулируя рост новой кости. В результате
средний пациент постоянно прибавляет около 3 дюймов (7-8 см) к своему росту в
полгода.
Сегодня в Китае растет беспокойство среди многих высокомобильных средних компаний.
класс мужчин и женщин, что они слишком короткие. Тысячи искали
решение этой проблемы путем удлинения ног. Это
достигается долгим и болезненным процессом, включающим хирургическое разрушение
две кости голени в обеих ногах, а затем с помощью регулируемых металлических скоб, которые
закреплены стальными штифтами, имплантированными в кость чуть ниже колен и
над лодыжками, чтобы постепенно увеличить длину почти на 1/16 дюйма
(около 1 мм.) в сутки по мере срастания костей. Это увеличивает разрыв в
зоны перелома, тем самым стимулируя рост новой кости. В результате
средний пациент постоянно прибавляет около 3 дюймов (7-8 см) к своему росту в
полгода.
Преднамеренная деформация тела сегодня является обычной практикой в Северной Америке.
Что ж. Для родителей из среднего и высшего класса принято иметь
зубы своих детей выпрямляли брекетами и брекетами. Это длинный,
дорогостоящий и несколько болезненный опыт, который изменяет положение зубов. Отчасти это делается для
сохранить и улучшить их работу. Однако сильная
мотивация – улучшить внешний вид. Выпрямление носа и др.
формы пластической хирургии часто делаются по той же причине, несмотря на
дело в том, что они болезненны. Родители предполагают, что эти виды
тела
изменение повысит вероятность того, что их дети вырастут более успешными в жизни. Это было
также мотивация богатых китайских родителей в прошлом, которые связывали ноги своим
дочерей и современных китайцев, которым делают удлинение ног.
Отчасти это делается для
сохранить и улучшить их работу. Однако сильная
мотивация – улучшить внешний вид. Выпрямление носа и др.
формы пластической хирургии часто делаются по той же причине, несмотря на
дело в том, что они болезненны. Родители предполагают, что эти виды
тела
изменение повысит вероятность того, что их дети вырастут более успешными в жизни. Это было
также мотивация богатых китайских родителей в прошлом, которые связывали ноги своим
дочерей и современных китайцев, которым делают удлинение ног.
Модная одежда |
Постоянный
изменения формы частей тела могут быть непреднамеренными. Например, носить кожу
обувь, закрывающая ступни, делает их уже, чем они были бы в противном случае. Точно так же практика ношения женщинами обуви с острыми носами, высоких каблуков и т.
часто слишком маленький размер обычно приводит к ряду болезненных ортопедических
деформации. Движущей силой этой внешне нелогичной западной культурной практики является
Вера в то, что маленькие ноги привлекательны для женщин.
Американская академия хирургов-ортопедов сообщила, что 9из 10 женщин
в Соединенных Штатах носят обувь, которая слишком мала для их ног, и 7 из
из десяти впоследствии развились болезненные косточки, молоткообразные пальцы или другие
деформации.
Например, носить кожу
обувь, закрывающая ступни, делает их уже, чем они были бы в противном случае. Точно так же практика ношения женщинами обуви с острыми носами, высоких каблуков и т.
часто слишком маленький размер обычно приводит к ряду болезненных ортопедических
деформации. Движущей силой этой внешне нелогичной западной культурной практики является
Вера в то, что маленькие ноги привлекательны для женщин.
Американская академия хирургов-ортопедов сообщила, что 9из 10 женщин
в Соединенных Штатах носят обувь, которая слишком мала для их ног, и 7 из
из десяти впоследствии развились болезненные косточки, молоткообразные пальцы или другие
деформации.
Что делает
такая корректировка развития частей нашего тела возможна благодаря тому факту, что люди
обладают высокой степенью физиологической пластичности . То есть мы можем быть
физически формируется окружающей средой в процессе выращивания. Взрослые
результат генетически унаследованных черт, которые в той или иной степени сформировались в каждом из нас
нашей средой, когда мы выросли.
| Ребенок с маразмом |
Крайнее недоедание или
переедание
в раннем детстве может привести к разрушительным изменениям в развитии.
Когда есть длительная нехватка пищи, как это бывает во время голода.
ситуация, у людей может развиться маразм (от греческого означает «чахнуть»). Симптомы включают экстремальные
истощение, диарея, анемия и апатия. Женщины с маразмом обычно
прекращают овуляцию и впоследствии не могут забеременеть. Потеря изолирующих жировых отложений делает людей с маразмом очень уязвимыми перед смертью в результате падения
основное тело
температура, когда воздух падает ниже 60-65 F.
(15-18 в.). Молодой
дети, пережившие маразм, обычно заканчивают
невысокий взрослый рост и некоторая степень умственной отсталости.
К сожалению, маразм сегодня не редкость во всем мире. Приблизительно 31 миллион детей ежегодно умирают от недоедания, а 178
миллионов остановились в росте. Сейчас около 1 миллиарда человек
недоедают, и по сути столько же переедают и страдают
проблемы со здоровьем, связанные с ожирением.
Приблизительно 31 миллион детей ежегодно умирают от недоедания, а 178
миллионов остановились в росте. Сейчас около 1 миллиарда человек
недоедают, и по сути столько же переедают и страдают
проблемы со здоровьем, связанные с ожирением.
Ребенок с квашиоркором |
Недостаток определенных видов питательных веществ может
привести к другим опасным для жизни проблемам со здоровьем. Например, когда
у младенцев и очень маленьких детей диета с чрезвычайно низким содержанием белка , они, вероятно,
разработка квашиоркор . Также
Способствует этому состоянию недостаточное потребление витаминов А и Е, а также
как минералы цинк и селен. Типичные симптомы
квашиоркора включают отек (или
отек) из-за задержки жидкости (особенно в животе), палочкообразные ноги
и руки с небольшой жировой или мышечной массой, апатией и потерей волос и кожи
пигментация пятнами. Как и в случае с маразмом, дети с
квашиоркор, вероятно, будут апатичными и
иммунной системы, что снижает их способность бороться с инфекциями. Если
ребенок переживает квашиоркор, у него, вероятно, задержка роста.
Как и в случае с маразмом, дети с
квашиоркор, вероятно, будут апатичными и
иммунной системы, что снижает их способность бороться с инфекциями. Если
ребенок переживает квашиоркор, у него, вероятно, задержка роста.
Даже конкретный
дефицит витаминов сам по себе может привести к серьезным проблемам со здоровьем
для детей, несмотря на адекватное питание. Например, отсутствие
витамин D может вызвать заболевание костей, известное как рахит,
в то время как недостаточное количество витамина А может
вызвать необратимую слепоту и
нарушать иммунную систему. Почти 100 миллионов
люди в мире сегодня испытывают нехватку витамина А. Большинство живет в Азии.
Чтобы уменьшить этот дефицит, новый сорт генетически модифицированного риса
(«золотой рис») с относительно высоким содержанием витамина А теперь выращивают
широко в Азии. Однако диета, содержащая слишком много витамина А,
одинаково вредны. Это может вызвать врожденные дефекты (особенно расщепление неба)
и может мешать клеткам, которые производят новую кость, что приводит к
резко возрастает риск переломов.
| Современный высокий японский |
Развитие
корректировка приводит не только к дефектам и нарушениям. Диетические изменения также могут иметь
положительный эффект, если питание
улучшенный. Так было в Японии после окончания мировой войны.
II. Министерство образования Японии сообщило, что
с тех пор в каждом поколении дети становились значительно выше. В
1986, например, 14-15-летние японские мальчики были в среднем на 7 дюймов выше, чем
мальчики сопоставимого возраста в 1959 году. Ключевой фактор изменения
в японском образе жизни была диета. Вполне вероятно, что это было
в основном отвечает за увеличение размеров тела. Между 1961 г.
и 1971 г., потребление животного белка в Японии.
выросло на 37%, а потребление растительной пищи снизилось на 3%. В городах Японии
и других все более богатых районах Восточной Азии, пищевые привычки изменились
резко за последние несколько десятилетий.
Гамбургеры, пицца, жареный цыпленок и другие западные продукты с высоким содержанием жира очень популярны.
с молодыми и богатыми. В Японии сегодня около
четверть потребляемых калорий составляют жиры — это в 5 раз больше
чем сразу после ВОВ. Поддержка гипотезы
что изменения в диете такого рода могут привести к значительному развитию
корректировки являются недавними
двухлетнее исследование детей в Кении. Было установлено, что включение
всего 60 граммов (около двух ложек) мяса в день в рацион молодняка.
детей привело к развитию мышц плеча на 80% больше.
по сравнению с детьми, которые были строгими вегетарианцами. Диета, которая включала
сопоставимое количество молока вместо мяса привело к увеличению на 40%.
Продукты животного происхождения важны в рационе детей раннего возраста, потому что
они содержат питательные вещества, которые трудно получить из не мясных или немолочных продуктов
источники.
В городах Японии
и других все более богатых районах Восточной Азии, пищевые привычки изменились
резко за последние несколько десятилетий.
Гамбургеры, пицца, жареный цыпленок и другие западные продукты с высоким содержанием жира очень популярны.
с молодыми и богатыми. В Японии сегодня около
четверть потребляемых калорий составляют жиры — это в 5 раз больше
чем сразу после ВОВ. Поддержка гипотезы
что изменения в диете такого рода могут привести к значительному развитию
корректировки являются недавними
двухлетнее исследование детей в Кении. Было установлено, что включение
всего 60 граммов (около двух ложек) мяса в день в рацион молодняка.
детей привело к развитию мышц плеча на 80% больше.
по сравнению с детьми, которые были строгими вегетарианцами. Диета, которая включала
сопоставимое количество молока вместо мяса привело к увеличению на 40%.
Продукты животного происхождения важны в рационе детей раннего возраста, потому что
они содержат питательные вещества, которые трудно получить из не мясных или немолочных продуктов
источники. Однако чрезмерное употребление животных белков и жиров может привести к
ожирение и другие риски для здоровья.
Однако чрезмерное употребление животных белков и жиров может привести к
ожирение и другие риски для здоровья.
Акклиматизация
Все остальные формы приспособления к экологическим стрессам обычно обратимы независимо от того, они происходят в детства или взрослой жизни. Эти обратимые изменения упоминается как акклиматизация или адаптация . Полезно рассмотреть разные формы акклиматизации по продолжительности что они могут произойти.
| Формы Акклиматизация |
| Дубление является общей формой сезонного акклиматизационного |
Ан
пример длительной акклиматизации
люди, которые теряют лишний жир и очень стройны
в результате умеренного, длительного недоедания. Если они позже увеличат свой рацион до постоянного уровня
избыточных калорий, они, скорее всего, сохранят больше жира и в конечном итоге станут
тучный. Они проходят длительную акклиматизацию, когда изначально теряют жировые отложения.
и снова позже, когда они сохранят его. В обоих случаях они акклиматизируются.
имеющийся запас продуктов питания.
Если они позже увеличат свой рацион до постоянного уровня
избыточных калорий, они, скорее всего, сохранят больше жира и в конечном итоге станут
тучный. Они проходят длительную акклиматизацию, когда изначально теряют жировые отложения.
и снова позже, когда они сохранят его. В обоих случаях они акклиматизируются.
имеющийся запас продуктов питания.
Анатомический и/или физиологические приспособления также могут развиваться в течение еще более коротких периодов времени. Например, многие люди приобретают темный загар в летние месяцы и теряют его. зимой. Это изменение окраски кожи является сезонной акклиматизацией. к разрушительному действию ультрафиолетового излучения солнца.
Когда кожа
водолазы погружаются в океан, они испытывают резко возрастающее давление воды.
Через несколько секунд они могут страдать от мучительной боли в ушах из-за неравномерного
давление внутри и снаружи барабанных перепонок. Они должны уравнять это давление путем
сильно дуют через нос. Делая это, они делают краткосрочный
акклиматизация к изменившейся среде.
Делая это, они делают краткосрочный
акклиматизация к изменившейся среде.
|
Когда
путешествуя на большую высоту, часто наблюдается прогрессирующее падение
способность слышать из-за неравенства давления с одной стороны
барабанная перепонка к другой, из-за чего она слегка раздувается
и стать менее гибким. Эта разница в давлении
опыт работы в горах обычно может быть
компенсируется зеванием, глотанием или жеванием жвачки. Однако, если
у кого-то заложен нос, часто трудно выровнять давление
когда они поднимаются на большую высоту.
В результате они могут испытывать сильную боль в ушах.
Это было серьезной проблемой для летчиков до того времени, когда самолеты
под давлением.
Различие между видами акклиматизации не только в количество времени, необходимое для первоначальной адаптации. Обычно, чем короче время для акклиматизации, тем быстрее он и в обратном направлении, как только окружающая среда стресса больше нет.
Комбинированные эффекты
Генетический
адаптация и три типа адаптации к стрессам окружающей среды не всегда
отдельные явления. Акклиматизация, происходящая в детстве, может привести к стойкому
анатомические изменения, как это часто бывает при недоедании. Когда проходит акклиматизация.
успешен в обеспечении хорошего здоровья и долголетия, он может дать людям избирательный
преимущество в передаче своих генов следующему поколению. Это может иметь сильное
определяющее влияние на направление эволюции. В свою очередь, генетические изменения могут сыграть
важную роль в адаптации, поскольку способность к акклиматизации в конечном итоге зависит от
генетический состав.
Адаптивность к конкретным экологическим стрессам варьируется от человека к человеку и от населения к Население. Мы не все биологически равны. Например, некоторые группы люди более успешно приспосабливаются к большой высоте. Другие могут лучше справиться сильная жара и высокая влажность. Адаптивные реакции, как правило, возникают в пространственные кластеры по всему миру. Как правило, наиболее эффективные приспособления для специфические экологические стрессы обнаруживаются в районах, где эти стрессы наиболее общий. Это свидетельствует о том, что естественный отбор имел место в успешно Адаптация населения.
Культура
Практики и технологии
Это
Важно помнить, что люди взаимодействуют не только с окружающей средой.
биологически. Мы также используем культуру. За последние полмиллиона лет
по крайней мере, мы
изобрели технологические средства, которые позволили нам занимать новые места без необходимости
сначала вырабатывают биологические приспособления к ним. Дома, одежда и огонь позволили нам жить
в умеренных и, в конечном счете, арктических регионах, несмотря на то, что мы все еще по существу имеем
тела тропических животных.
Дома, одежда и огонь позволили нам жить
в умеренных и, в конечном счете, арктических регионах, несмотря на то, что мы все еще по существу имеем
тела тропических животных.
Летчик-истребитель FA-18 |
Однако это не означает, что искусственная технология устраняет биологическое адаптивные преимущества отдельных лиц или групп. Люди, у которых толще слои жировой изоляции под их кожей все еще обычно лучше выживают в холодном климате, в то время как стройным людям лучше в горячих.
В следующих четырех разделах руководства вы узнаете, как наши тела реагируют на несколько распространенных видов стрессов окружающей среды.
| Вернуться в меню | Практический тест | Следующий Тема |
Журнальная статья
М. Х. Песпени,
Х. Песпени,
М. Х. Песпени 1
Ищите другие работы этого автора на:
Оксфордский академический
пабмед
Google ученый
Ф. Чан,
Ф. Чан
Ищите другие работы этого автора на:
Оксфордский академический
пабмед
Google ученый
Б. А. Менге,
Б. А. Менге
Ищите другие работы этого автора на:
Оксфордский академический
пабмед
Google ученый
С. Р. Палумби
С. Р. Палумби
Ищите другие работы этого автора на:
Оксфордский академический
пабмед
Google ученый
Примечания автора
Интегративная и сравнительная биология , том 53, выпуск 5, ноябрь 2013 г. , страницы 857–870, https://doi.org/10.1093/icb/ict094
, страницы 857–870, https://doi.org/10.1093/icb/ict094
Опубликовано:
30004 9002 600
- Разделенный вид
- Содержание статьи
- Рисунки и таблицы
- видео
- Аудио
- Дополнительные данные
Цитировать
Процитировать
М.
 Х. Песпени, Ф. Чан, Б. А. Менге, С. Р. Палумби, Признаки адаптации к местным условиям рН в мозаике окружающей среды в экосистеме Калифорнийского течения, Интегративная и сравнительная биология , том 53, выпуск 5, ноябрь 2013 г., страницы 857–870, https://doi.org/10.1093/icb/ict094
Х. Песпени, Ф. Чан, Б. А. Менге, С. Р. Палумби, Признаки адаптации к местным условиям рН в мозаике окружающей среды в экосистеме Калифорнийского течения, Интегративная и сравнительная биология , том 53, выпуск 5, ноябрь 2013 г., страницы 857–870, https://doi.org/10.1093/icb/ict094Выберите формат Выберите format.ris (Mendeley, Papers, Zotero).enw (EndNote).bibtex (BibTex).txt (Medlars, RefWorks)
Закрыть
Разрешения
- Электронная почта
- Твиттер
- Подробнее
Фильтр поиска панели навигации Интегративная и сравнительная биологияЭтот выпускЖурналы SICBБиологические наукиКнигиЖурналыOxford Academic Термин поиска мобильного микросайта
Закрыть
Фильтр поиска панели навигации Интегративная и сравнительная биологияЭтот выпускЖурналы SICBБиологические наукиКнигиЖурналыOxford Academic Термин поиска на микросайте
Advanced Search
«> Abstract
Мало что известно о возможности быстрой эволюции естественных популяций в ответ на высокие темпы современных климатических изменений. Особый интерес представляют организмы, развившиеся в средах с высокой изменчивостью в пространстве и времени, поскольку они могут содержать генетические вариации, которые могут способствовать эволюционной реакции на меняющиеся условия. Здесь мы рассмотрим, что известно о генетической способности к адаптации пурпурного морского ежа, Strongylocentrotus purpuratus , вид, который эволюционировал в апвеллинговой экосистеме северо-восточной части Тихого океана. Мы также представляем новые результаты тестирования адаптации к местным условиям рН в шести популяциях от Орегона до южной Калифорнии. Мы объединяем данные о 19 493 генетических полиморфизмах с данными о местных условиях рН. Мы обнаружили корреляции между частотой аллелей и средним ранговым временем, проведенным при pH <7,8, в 318 однонуклеотидных полиморфизмах в 275 генах. Два гена, наиболее коррелирующие с локальным pH, представляют собой белок, связанный с цитоскелетом и протонным насосом, с функциональной ролью в поддержании объема клетки и внутренней регуляции pH соответственно.
Особый интерес представляют организмы, развившиеся в средах с высокой изменчивостью в пространстве и времени, поскольку они могут содержать генетические вариации, которые могут способствовать эволюционной реакции на меняющиеся условия. Здесь мы рассмотрим, что известно о генетической способности к адаптации пурпурного морского ежа, Strongylocentrotus purpuratus , вид, который эволюционировал в апвеллинговой экосистеме северо-восточной части Тихого океана. Мы также представляем новые результаты тестирования адаптации к местным условиям рН в шести популяциях от Орегона до южной Калифорнии. Мы объединяем данные о 19 493 генетических полиморфизмах с данными о местных условиях рН. Мы обнаружили корреляции между частотой аллелей и средним ранговым временем, проведенным при pH <7,8, в 318 однонуклеотидных полиморфизмах в 275 генах. Два гена, наиболее коррелирующие с локальным pH, представляют собой белок, связанный с цитоскелетом и протонным насосом, с функциональной ролью в поддержании объема клетки и внутренней регуляции pH соответственно. Во всех протестированных локусах высокие корреляции с локальным рН были сосредоточены в генах, связанных с транспортом ионов, биоминерализацией, метаболизмом липидов и межклеточной адгезией, функциональными путями, важными для поддержания гомеостаза при низком рН. Мы определили набор из семи генов в качестве главных кандидатов на быструю эволюционную реакцию на закисление океана. В этих генах предполагаемый адаптированный к низкому pH аллель, основанный на частотах аллелей в естественных популяциях, быстро увеличивается по частоте у личинок пурпурных морских ежей, выращенных при низком pH. Мы также обнаружили, что популяции из местностей с высоким pH показывают большее изменение частоты аллелей в сторону предполагаемых адаптированных к низкому pH аллелей при экспериментальном подкислении по сравнению с популяциями с низким pH, что свидетельствует о том, что как естественный, так и искусственный отбор отдают предпочтение одним и тем же аллелям для реакции на низкий рН. Эти результаты показывают, что пурпурные морские ежи могут быть адаптированы к местному pH, и предполагают, что этот вид может обладать генетической способностью к быстрой эволюции в ответ на закисление.
Во всех протестированных локусах высокие корреляции с локальным рН были сосредоточены в генах, связанных с транспортом ионов, биоминерализацией, метаболизмом липидов и межклеточной адгезией, функциональными путями, важными для поддержания гомеостаза при низком рН. Мы определили набор из семи генов в качестве главных кандидатов на быструю эволюционную реакцию на закисление океана. В этих генах предполагаемый адаптированный к низкому pH аллель, основанный на частотах аллелей в естественных популяциях, быстро увеличивается по частоте у личинок пурпурных морских ежей, выращенных при низком pH. Мы также обнаружили, что популяции из местностей с высоким pH показывают большее изменение частоты аллелей в сторону предполагаемых адаптированных к низкому pH аллелей при экспериментальном подкислении по сравнению с популяциями с низким pH, что свидетельствует о том, что как естественный, так и искусственный отбор отдают предпочтение одним и тем же аллелям для реакции на низкий рН. Эти результаты показывают, что пурпурные морские ежи могут быть адаптированы к местному pH, и предполагают, что этот вид может обладать генетической способностью к быстрой эволюции в ответ на закисление. Эта адаптивная способность, вероятно, возникает из-за постоянной генетической изменчивости, поддерживаемой в природе путем уравновешивания отбора в пространственной и временной мозаике окружающей среды, которая характеризует Калифорнийскую текущую экосистему.
Эта адаптивная способность, вероятно, возникает из-за постоянной генетической изменчивости, поддерживаемой в природе путем уравновешивания отбора в пространственной и временной мозаике окружающей среды, которая характеризует Калифорнийскую текущую экосистему.
Здесь мы сосредоточимся на пурпурном морском еже, Strongylocentrotus purpuratus , как на модельном организме для понимания адаптивного генетического ландшафта для видов с высокой дисперсией в мозаике окружающей среды. Пурпурный морской еж — это широко распространенное нерестящееся морское беспозвоночное, которое можно найти в большинстве приливных и сублиторальных скалистых рифов от холодных вод Аляски до более теплых вод Нижней Калифорнии, Мексика (Rogers-Bennett 2007). Геномные инструменты, понимание экологии и усиление мониторинга окружающей среды, связанное с пурпурными морскими ежами, дают важные сведения о том, как изменчивость окружающей среды и эволюционная сила уравновешивающего отбора могут поддерживать адаптивную генетическую изменчивость в популяциях пурпурных морских ежей (Консорциум по секвенированию генома морских ежей и др. , 2006; Песпени и др., 2012; Эванс и др., 2013; Келли и др., 2013; Песпени и Палумби, 2013). Этот резервуар адаптивных аллелей может иметь решающее значение для эволюционных реакций пурпурных ежей и других морских видов на будущие климатические изменения. В результате понимание способности и пределов постоянной генетической изменчивости для подпитки будущих адаптивных сдвигов для этого вида может быть иллюстрацией других видов в восточной части Тихого океана и в других местах.
Эта статья состоит из трех основных разделов. Во-первых, мы рассмотрим недавние эволюционные исследования биологии популяции пурпурного морского ежа. Во-вторых, мы рассматриваем доказательства эволюции регуляции генов и роли пространственного и временного уравновешивающего отбора, позволяющего этому виду сохраняться в мозаике окружающей среды. В-третьих, мы представляем новые результаты ранее опубликованных данных, которые демонстрируют связь между локальным pH и генетической изменчивостью. Наша предыдущая работа показала, как низкий pH был фактором отбора множества генов в геномах личинок пурпурных морских ежей (Pespeni et al. 2013b). Текущая работа дополняет экологический контекст этих результатов и предоставляет ключевые гены-кандидаты и пути, которые могут обеспечить устойчивость у этого и других видов. Мы исследуем естественные различия между популяциями в степени их адаптации к местному pH и реакции на экспериментальное подкисление.
Несмотря на широкую неоднородность окружающей среды вдоль ареала вида, предыдущие исследования с использованием митохондриальных и аллозимных маркеров практически не выявили структуры популяций вдоль ареала вида (Палумби и Уилсон 19).90; Эдмандс и др. 1996 год; Песпени и Палумби, 2013 г.). Полногеномное сканирование 12 431 однонуклеотидного полиморфизма (SNP) показывает, что все аллели в этих участках присутствуют в двух популяциях, разделенных более чем 1700 км (Бойлер-Бей, Орегон, и Сан-Диего, Калифорния). То есть нет фиксированных различий в аллелях между этими популяциями на этих 12 431 участках генома (Pespeni et al. 2010). Отсутствие прочной генетической структуры и совместное использование аллелей в пространстве согласуются с высоким потенциалом морских ежей к расселению в результате внешнего оплодотворения и стадии жизненного цикла плавающих и питающихся личинок (Strathmann 19).78).
Несмотря на то, что среди популяций пурпурных морских ежей существует высокий потенциал расселения и общей генетической однородности, есть несколько признаков ограниченного потока генов в определенных генах по всему геному, что свидетельствует о локальной адаптации. Хотя между морскими ежами с побережья Орегона и Калифорнии нет фиксированных аллельных различий, полногеномные паттерны частот аллелей достаточно различаются, чтобы можно было различать популяции с помощью анализа основных компонентов, разделяя генетическую изменчивость в пространстве (рис. 1А) (Pespeni). и др., 2010). Анализы, объединяющие данные о функциях генов, определяют определенные классы генов, которые могут быть важны для местной адаптации; в частности, гены в пути деградации белка Ubiquitin демонстрируют высокие уровни генетической дифференциации по сравнению со средним значением для всего генома (Pespeni et al. 2012). Дифференцированные гены Ubiquitin представляют собой лигазы E3, мишень-специфические регуляторы нескольких клеточных процессов от метаболизма до развития и запрограммированной гибели клеток (Pespeni et al. 2012). Кроме того, существует избыток аллельного разнообразия в генах, связанных с иммунитетом, в южном населении, где заболеваемость высока, что позволяет предположить, что существует селективное давление в пользу большего разнообразия в иммунных генах, подобное тому, что наблюдалось в главном комплексе гистосовместимости. MHC) локусов в адаптивных иммунных системах позвоночных (рис. 1В) (Pespeni et al. 2012). Взятые вместе, эти результаты предполагают, что естественный отбор действовал на определенные гены по всему геному, оставляя признаки уменьшения потока генов в одних классах генов и повышения генетического разнообразия в других. Эти сложные паттерны геномного разнообразия могут быть возможны благодаря высокому уровню генетического разнообразия и высокой скорости рекомбинации у этого вида, а также эволюционным процессам, таким как уравновешивающий отбор и плавный отбор (Хермисон и Пеннингс, 2005; Песпени и Палумби, 2013). .
Рис. 1
Открыть в новой вкладкеСкачать слайд
Дифференциация популяций и признаки локальной адаптации у широко расселенного пурпурного морского ежа. Образцы были взяты из Бойлер-Бей, штат Орегон (север) и Сан-Диего, штат Калифорния (юг). ( A ) Разделение популяций на основе разделения дисперсии в генетической изменчивости 12 431 полногеномного полиморфизма с использованием анализа основных компонентов. Адаптировано из Pespeni et al. (2010). ( B ) Признаки уравновешивающего отбора в повышенной гетерозиготности в кодирующих областях генов, связанных с иммунитетом. В то время как уровни гетерозиготности по всем локусам выше у южных ежей, гетерозиготность по генам иммунитета непропорционально выше в южных популяциях, где давление болезней выше, и, таким образом, наводит на мысль о местной адаптации. Адаптировано из Pespeni et al. (2012).
Последующее исследование предполагаемых адаптивных локусов предполагает, что сочетание естественного отбора и локального сохранения аллелей в районе Южно-Калифорнийского залива может позволить популяциям пурпурных морских ежей адаптироваться к особым условиям окружающей среды, характерным для этого региона (Pespeni и Палумби, 2013 г. ). Семь генов были идентифицированы как предположительно адаптивные или предположительно нейтральные на основании исследования сканирования генома (Pespeni et al. 2010). Эти гены были изучены более подробно путем прямого секвенирования гораздо большего количества особей, отобранных из шести популяций, охватывающих большую часть ареала вида от Бэмфилда, Канада, до Пунта-Сан-Карлос, Мексика (Pespeni and Palumbi 2013). Предполагаемые адаптивные локусы демонстрировали признаки отбора, в то время как предполагаемые нейтральные локусы обычно этого не делали (более подробно обсуждается в следующем разделе). Паттерны во всех пяти предполагаемых адаптивных локусах показали снижение потока генов в популяцию Сан-Диего, расположенную в Южно-Калифорнийском заливе, и из нее; паттерн, не наблюдаемый в предполагаемых нейтральных генах (Pespeni and Palumbi 2013). Южная Калифорнийская бухта имеет более теплые воды, поскольку прибрежные рифы расположены у берега широкого континентального шельфа, защищенного от апвеллинга, и отличаются от более холодных, апвеллинговых береговых линий на севере и юге (Бакун и Пэрриш 19). 82; Смит и Эппли, 1982 г.; Зайцев и др. 2003 г.; Манн и Лазиер, 2006). Признак дифференцировки в Сан-Диего предполагает, что может действовать адаптация к местной температуре, pH или заболеванию. В этом регионе масштаб изменчивости окружающей среды может быть больше, чем масштаб расселения личинок, что позволяет накапливать разные аллели в нескольких поколениях.
Хотя эти исследования выявили предполагаемое функциональное разнообразие в регуляторных областях генов, еще предстоит определить, существуют ли различия в регуляции генов между популяциями. Песпени и др. (2013a) провели первое долгосрочное исследование акклиматизации взрослых пурпурных морских ежей из отдаленных популяций в обычном саду (Pespeni et al. 2013a). Цель акклиматизации в обычном саду состоит в том, чтобы стереть различия в истории окружающей среды, чтобы выявить генетически контролируемые различия, в данном случае различия в регуляции генов (Prosser 19). 86; Хочачка и Сомеро 2002). Песпени и др. выращивали морских ежей из Сан-Диего, штат Калифорния, и Бойлер-Бей, штат Орегон, вместе на морской станции Хопкинса в Монтерее, Калифорния, в течение 3 лет, а затем взяли образцы тканей этих совместно культивируемых особей для оценки паттернов экспрессии генов в масштабах всего транскриптома. Авторы измерили уровни распространенности транскриптов 18 883 генов с помощью РНК-секвенирования. В то время как большая часть транскриптома показала одинаковые уровни экспрессии генов между отобранными популяциями, исследование выявило стойкие различия в регуляции генов, связанных с биоминерализацией и ростом (Pespeni et al. 2013a).
Экспрессия генов, связанных с биоминерализацией и метаболизмом, была постоянно выше у южных ежей по сравнению с северными ежами (рис. 2). Различия предсказывают, что у южных ежей будет больше возможностей для роста, чем у северных ежей в условиях обычного сада. Апостериорный эксперимент подтвердил, что у южных ежей действительно отрастали иглы быстрее, чем у северных ежей (Pespeni et al. 2013a). Эти результаты продемонстрировали, что фенотип экспрессии генов первого порядка по всему транскриптому имеет морфологические последствия. Более высокие возможности для роста южных ежей могут указывать на то, что ежи, обитающие в районе Южно-Калифорнийского залива, процветают в более прохладных, богатых ресурсами условиях обычного сада Монтерей-Бей (ежи кормились ad libitum ) по сравнению с ежами, обитающими на побережье Орегона, которые испытали относительно более высокие температуры и более высокие уровни pH в обычном саду залива Монтерей. Эти генно-регуляторные различия, по-видимому, контролируются генетически и могут иметь последствия для адаптированных к местным условиям популяций морских ежей, поскольку они испытывают изменение климатических режимов. Изменения условий окружающей среды, влияющие на рост или биоминерализацию, такие как изменения температуры или рН, могут оказывать неравномерное воздействие на разные популяции.
Рис. 2
Открыть в новой вкладкеСкачать слайд
Стойкие различия в регуляции 10% генов биоминерализации между долгоживущими, акклиматизированными в обычных садах пурпурными морскими ежами из Бойлер-Бей, Орегон, и Сан-Диего, Калифорния. Каждая точка на точечной диаграмме представляет среднюю экспрессию генов у северных ежей по сравнению с южными ежами для 296 генов, связанных с биоминерализацией у пурпурных морских ежей; треугольники показывают гены, дифференциально экспрессируемые во всех 18 883 генах, проверенных на различия в экспрессии (FDR P < 0,05), ромбы добавляют гены, дифференциально экспрессируемые из подмножества биоминерализации ( q — значение < 0,05), в то время как незакрашенные кружки показывают гены, связанные с биоминерализацией, которые не экспрессировались дифференциально между популяциями (FDR P > 0,05 ). Диагональная линия представляет собой линию 1:1. Среди 10% дифференциально экспрессирующихся генов у южных ежей экспрессия выше, чем у северных, во всех случаях. Адаптировано из Pespeni et al. (2013а).
Эволюция регуляции генов и уравновешивающий отбор, вероятно, являются важными факторами в поддержании функционального разнообразия в экологически неоднородном морском ландшафте. Как описано в предыдущем разделе, Песпени и Палумби протестировали пять локусов, имевших высокую генетическую дифференциацию при предыдущем сканировании генома (Песпени и др., 2010), на наличие признаков естественного отбора на уровне последовательности, используя прямое секвенирование у пурпурных морских ежей из популяций, охватывающих большую часть ареала вида от Канады до Нижней Калифорнии, Мексика (Pespeni and Palumbi 2013). Предполагаемые адаптивные локусы показали признаки отбора, включая корреляции с температурой, а не с широтой, или избыток изменений в аминокислотах, в отличие от протестированных предполагаемых нейтральных локусов. Два из пяти протестированных предполагаемых адаптивных локусов были транскрипционными факторами. Оба этих гена демонстрируют высокие уровни полиморфизма, изменяющего аминокислоты, сохраняющиеся в шести исследованных популяциях (dN/dS >> 1), а также сигналы пространственного или временного балансирующего отбора (см. рис. 3A и B для частот аллелей в популяциях). ) (Песпени и Палумби, 2013). Многие аллели в этих генах факторов транскрипции различаются последовательностями аминокислот. Функция этих факторов транскрипции либо особенно невосприимчива к вариациям аминокислот, либо существует большое количество функциональных вариаций среди людей в генах, регулируемых этими факторами транскрипции.
В другом исследовании изучалась эволюция регуляции генов у пурпурных морских ежей, чтобы обнаружить высокие уровни генетической изменчивости и сигналы положительного отбора в регуляторных областях шести из восьми протестированных генов (Garfield et al. 2012). Эти гены, участвующие в раннем развитии морских ежей, были выбраны для исследования, потому что их регуляторные области были хорошо охарактеризованы в предыдущих исследованиях, а не потому, что они были идентифицированы как предполагаемые цели отбора. Этот процесс отбора локуса делает сигналы естественного отбора еще более поразительными и предполагает, что пути регуляции генов, занимающие центральное место в раннем развитии, могут быть более чувствительными к давлениям отбора и менее каноническими, чем обычно считается. Также следует отметить, что высокие уровни генетического разнообразия и сигналы уравновешивающего отбора сохраняются в одной популяции, отобранной в Санта-Барбаре, Калифорния (Garfield et al. 2012). Таким образом, эти исследования подчеркивают роль уравновешивающего отбора в поддержании полиморфизмов, которые изменяют регуляцию генов внутри и между популяциями вида в неоднородном наземном или морском ландшафте, как подходящей области для дальнейших исследований, особенно в контексте быстро меняющихся климатических условий.
Рис. 3
Открыть в новой вкладкеСкачать слайд
Избыточная изменчивость аминокислот и сигналы пространственного или временного балансирующего отбора в двух белках-факторах транскрипции: факторе транскрипции 25 ( A , SPU_015723) и факторе транскрипции ( B , SPU_002852). Линии показывают частоты аллелей каждого SNP в популяциях с севера на юг, Бэмфилд, Канада (BM), Бойлер-Бей, Орегон (BB), Патрик-Пойнт, Калифорния (PP), Монтерей, Калифорния (MY), Сан-Диего, Калифорния (SD). ) и три участка, отобранные в Нижней Калифорнии, Мексика (Баха). RSTA (Restriction-Site Tiling Analysis) обозначает полиморфизмы, изначально идентифицированные как дифференцированные между популяциями. Адаптировано из Песпени и Палумби (2013 г.).
Подкисление океанов представляет серьезную угрозу для организмов, выделяющих скелеты и раковины из карбоната кальция, таких как кораллы, мидии, морские ежи и планктон (Harley et al. 2006; Hoegh-Guldberg et al. 2007; Doney et al. 2009 г.). Однако Калифорнийская текущая экосистема является отличной естественной лабораторией для изучения потенциала эволюционной адаптации к низкому pH. Региональные различия в сезонном подъеме холодных, богатых питательными веществами вод с низким pH вдоль побережья Орегона и Калифорнии создают пространственно и во времени изменчивую мозаику условий pH окружающей среды (Feely et al. 2008). Организмы, живущие в этой среде обитания, сезонно испытывают условия pH, подобные тем, которые были предсказаны для 2100 года (Hofmann et al. 2011).
На сегодняшний день несколько исследований продемонстрировали способность к эволюции в ответ на высокое содержание CO 2 ряда морских видов, включая три вида ежей, виды мшанок, виды мидий и несколько штаммов видов кокколитофорид (Langer et и др. , 2009 г.; Риджвелл и др., 2009 г.; Финдли и др., 2011 г.; Паркер и др., 2011 г.; Пистевос и др., 2011 г.; Сандей и др., 2011 г.; Фу и др., 2012 г.; Келли и др., 2013 г.; Песпени и др. 2013б). В этих исследованиях организмы из одной или нескольких природных популяций были доставлены в лабораторию и культивированы в условиях высокого содержания CO 9 .0181 2 . Эти эксперименты показали, что организмы имеют постоянную генетическую изменчивость для наследственных реакций на климатические изменения.
В настоящем исследовании мы выясняем, какие естественные популяции пурпурных морских ежей несут генетическую изменчивость для эволюционной реакции на низкий рН, путем проверки корреляций между локальными условиями рН, наблюдаемыми в природе, и локальными частотами аллелей . Мы также проверяем, являются ли предполагаемые аллели, адаптированные к низкому pH, идентифицированные в природных популяциях, аллелями, которые проявляют наибольшую реакцию на экспериментальное подкисление. Если популяции адаптируются к местным условиям pH, мы ожидаем, что подтвердятся два простых предсказания: (1) популяции, испытывающие хронически высокие уровни CO 2 содержат аллели, улучшающие выживаемость в условиях высокого содержания CO 2 и (2) что эти аллели должны демонстрировать наибольшую реакцию на экспериментальное закисление в популяциях, которые не испытывают хронического высокого содержания CO 2 . Чтобы подтвердить первое предсказание, мы ожидаем найти больше аллелей, коррелирующих с локальным pH, чем ожидалось случайно, и что эти коррелированные аллели будут в генах, которые, по прогнозам, будут играть важную роль в поддержании гомеостаза при высоком уровне CO 9 .0181 2 условия. Чтобы подтвердить второе предсказание, мы ожидаем, что личинки из популяций с низким содержанием CO 2 в природе будут демонстрировать большие изменения частоты аллелей в ответ на экспериментальное закисление в предполагаемых адаптированных генах с высоким содержанием CO 2 .
Таблица 1
Места отбора проб, сокращения, координаты и местные условия рН для шести мест сбора взрослых ежей
| Местоположение . | Аббревиатура сайта. . | Широта . | Долгота . | Уровень кислотности
. | Среднее значение рН . | SD рН . |
|---|---|---|---|---|---|---|
| Fogarty Creek, OR | FC | 44.81 | 124.1 | 1 | 7.97 | 0.18 |
| Strawberry Hill, OR | ST | 44.25 | 124.1 | 2 | 8.02 | 0.17 |
| Van Damme State Park, CA | VD | 39.28 | 123.8 | 4 | 7.96 | 0.09 |
| Bodega Marine Reserve, CA | BD | 38.31 | 123.1 | 3 | 7.99 | 0.14 |
| Terrace Point, CA | TP | 36.95 | 122.1 | 6 | 8.10 | 0.11 |
| Alegria, CA | AL | 34.25 | 119,6 | 5 | 8,08 | 0,13 |
9
0999999999999999999999999999999999999999999999999999999999999999999999999999999999999999999999999н
.
11 Rank acidity was determined by the amount of time spent при pH < 7,8 на каждом участке с апреля по сентябрь 2011 г., период времени, охватывающий сезон апвеллинга для всех участков.
Открыть в новой вкладке
Таблица 1
Места отбора проб, сокращения, координаты и местные условия pH для шести мест сбора взрослых ежей
| Местоположение . | Аббревиатура сайта. . | Широта . | Долгота . | Уровень кислотности . | Среднее значение рН . | SD рН . |
|---|---|---|---|---|---|---|
| Фогерти-Крик, Орегон | FC | 44.81 | 124.1 | 1 | 7. 97 | 0.18 |
| Strawberry Hill, OR | ST | 44.25 | 124.1 | 2 | 8.02 | 0.17 |
| Van Damme Государственный парк, CA | VD | 39.28 | 123,8 | 4 | 7,96 | 0,09 |
| Bodega Marine Reserve, CA | 4 | |||||
| Bodega Marine, CA | 9 | |||||
| .0331 | 38.31 | 123.1 | 3 | 7.99 | 0.14 | |
| Terrace Point, CA | TP | 36.95 | 122.1 | 6 | 8.10 | 0.11 |
| Alegria, CA | AL | 34.25 | 119,6 | 5 | 8,08 | 0,13 |
| Аббревиатура сайта. . | Широта . | Долгота
. | Уровень кислотности . | Среднее значение рН . | SD рН . | |
|---|---|---|---|---|---|---|
| Fogarty Creek, OR | FC | 44.81 | 124.1 | 1 | 7.97 | 0.18 |
| Strawberry Hill, OR | ST | 44.25 | 124.1 | 2 | 8.02 | 0.17 |
| Van Damme State Park, CA | VD | 39.28 | 123.8 | 4 | 7.96 | 0.09 |
| Bodega Marine Reserve, CA | BD | 38.31 | 123.1 | 3 | 7.99 | 0.14 |
| Terrace Point, CA | TP | 36.95 | 122.1 | 6 | 8.10 | 0.11 |
| Alegria, CA | AL | 34.25 | 119.6 | 5 | 8. 08 | 0.13 |
Rank acidity was determined by the amount of time spent при pH < 7,8 на каждом участке с апреля по сентябрь 2011 г., период времени, охватывающий сезон апвеллинга для всех участков.
Открыть в новой вкладке
Мы идентифицировали гены, потенциально адаптированные к местным условиям рН, путем проверки корреляции между частотой аллелей в популяции и ранговым локальным рН. Мы идентифицировали 318 SNP, присутствующих в 275 генах, коррелирующих с локальным рН ( P < 0,01), на 65% больше, чем ожидалось случайно из 19 493 протестированных SNP. Было 1439 SNP, коррелирующих с локальным pH на уровне P < 0,05, что на 47% больше, чем ожидалось случайно, хотя ни один набор не выдержал поправки на частоту ложных открытий (FDR P > 0,05). Топ-10 аннотированных генов с коррелирующими SNP перечислены в таблице 2. Они включают белок цитоскелета, белок протонной помпы и белок-носитель стерола. стабильность при низком pH (Putnam 1998; Сази и др. 2001 г.; Пёртнер 2008). См. Дополнительную таблицу S1 для получения полного списка из 318 SNP с частотами популяционных аллелей. Эти результаты предполагают действие естественного отбора в ответ на местные условия pH, поскольку существует больше SNP, чем ожидалось случайно, которые коррелировали с pH. Более того, идентифицированные SNP находятся в генах, которые, по прогнозам, важны для физиологического ответа на низкий pH.
Таблица 2
Топ-10 генов с частотами аллелей в природных популяциях, коррелирующими с локальными условиями pH (из 318 SNP с P < 0,01)
| Генная аннотация . | Идентификатор гена . | SNP позиция . | Ср. нет. считывает на образец . | Корреляция ( r ) . | P -значение . |
|---|---|---|---|---|---|
| Белок, ассоциированный с цитоскелетом | SPU_004863 | 261 | 1254 | 0. 96 | 0.001 |
| Glutamic-oxaloacetic transaminase 2 | SPU_014067 | 372 | 1201 | 0.82 | 0.002 |
| Vacuolar proton pump delta polypeptide | SPU_012628 | 192 | 817 | 0,81 | 0,002 |
| Лизил-тРНК-синтетаза | SPU_008981 | 522 | 1057 1 | 0,8303310.002 | |
| Immunomodulatory protein | SPU_004320 | 239 | 572 | 0.81 | 0.001 |
| Endoplasmic reticulum-golgi intermediate | SPU_017463 | 270 | 1043 | 0.80 | 0.002 |
| Белок-носитель стерола 2 изоформа 1 пропротеин | SPU_006383 | 432 | 830 | 0,78 | 0.002 |
| Hydroxyacyl-coenzyme A | SPU_008554 | 768 | 1614 | 0. 76 | 0.002 |
| Frizzled homolog 8a | SPU_022916 | 945 | 935 | 0.76 | 0.001 |
| Эукариотический фактор инициации трансляции 3 | SPU_026863 | 1281 | 1505 | 0,74 | 0,002 | 4
| Генная аннотация . | Идентификатор гена . | SNP позиция . | Ср. нет. считывает на образец . | Корреляция ( r ) . | P -значение . | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Белок, ассоциированный с цитоскелетом | SPU_004863 | 261 | 1254 | 0,96 | 0.001 | ||||||
| Glutamic-oxaloacetic transaminase 2 | SPU_014067 | 372 | 1201 | 0.82 | 0.002 | ||||||
| Vacuolar proton pump delta polypeptide | SPU_012628 | 192 | 817 | 0. 81 | 0,002 | ||||||
| Лизил-тРНК-синтетаза | SPU_008981 | 522 | 1057 | 0,81 | 0,81 0,03310331|||||||
| Immunomodulatory protein | SPU_004320 | 239 | 572 | 0.81 | 0.001 | ||||||
| Endoplasmic reticulum-golgi intermediate | SPU_017463 | 270 | 1043 | 0.80 | 0.002 | ||||||
| Sterol белок-носитель 2 изоформа 1 пробелок | SPU_006383 | 432 | 830 | 0,78 | 0,002 | ||||||
| Hydroxyacyl-coenzyme A | SPU_008554 | 768 | 1614 | 0.76 | 0.002 | ||||||
| Frizzled homolog 8a | SPU_022916 | 945 | 935 | 0.76 | 0.001 | ||||||
| Eukaryotic translation инициирующий фактор 3 | SPU_026863 | 1281 | 1505 | 0,74 | 0,002 |
| Генная аннотация . | Идентификатор гена . | СНиП позиция . | Ср. нет. считывает на образец . | Корреляция ( r ) . | P -значение . | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Cytoskeleton associated protein | SPU_004863 | 261 | 1254 | 0.96 | 0.001 | |||||
| Glutamic-oxaloacetic transaminase 2 | SPU_014067 | 372 | 1201 | 0.82 | 0. 002 | |||||
| Vacuolar proton pump delta polypeptide | SPU_012628 | 192 | 817 | 0.81 | 0.002 | |||||
| Lysyl-tRNA synthetase | SPU_008981 | 522 | 1057 | 0,81 | 0,002 | |||||
| Иммуномодулирующий белок | SPU_004320 | 239 | 8,1330 9031 | 0331 | 0.001 | |||||
| Endoplasmic reticulum-golgi intermediate | SPU_017463 | 270 | 1043 | 0.80 | 0.002 | |||||
| Sterol carrier protein 2 isoform 1 proprotein | SPU_006383 | 432 | 830 | 0.78 | 0,002 | |||||
| Гидроксиацил-кофермент А | SPU_008554 | 768 | 1614 | 0,7330 0,730330 0.002 | ||||||
| Frizzled homolog 8a | SPU_022916 | 945 | 935 | 0. 76 | 0.001 | |||||
| Eukaryotic translation initiation factor 3 | SPU_026863 | 1281 | 1505 | 0.74 | 0.002 |
| Генная аннотация . | Идентификатор гена . | SNP позиция . | Ср. нет. считывает на образец . | Корреляция ( r ) . | P -значение . | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Cytoskeleton associated protein | SPU_004863 | 261 | 1254 | 0.96 | 0.001 | ||||||
| Glutamic-oxaloacetic transaminase 2 | SPU_014067 | 372 | 1201 | 0.82 | 0.002 | ||||||
| Vacuolar proton pump delta polypeptide | SPU_012628 | 192 | 817 | 0.81 | 0. 002 | ||||||
| Lysyl-tRNA synthetase | SPU_008981 | 522 | 1057 | 0.81 | 0.002 | ||||||
| Иммуномодулирующий белок | SPU_004320 | 239 | 572 | 0,81 | 0,0011 | 1Endoplasmic reticulum-golgi intermediate | SPU_017463 | 270 | 1043 | 0.80 | 0.002 |
| Sterol carrier protein 2 isoform 1 proprotein | SPU_006383 | 432 | 830 | 0.78 | 0.002 | ||||||
| Гидроксиацил-кофермент А | SPU_008554 | 768 | 1614 | 0,76 | 0,002 | SPU_022916 | 945 | 935 | 0.76 | 0.001 | |
| Eukaryotic translation initiation factor 3 | SPU_026863 | 1281 | 1505 | 0.74 | 0. 002 |
For additional candidates, see Дополнительная таблица S1 для полного списка из 318 SNP, коррелирующих с локальными данными о pH и частоте аллелей сайта.
Открыть в новой вкладке
2013b). Действительно, окисление липидов, катаболизм жирных кислот и анионный транспорт входят в число девяти основных категорий (9).0014 P < 0,01). В целом, восемь из девяти категорий, обогащенных высокой корреляцией с локальным pH, играют физиологические роли, которые, как ожидается, на основе предыдущей работы будут участвовать в поддержании гомеостаза при низком pH (Pörtner et al., 2005; Melzner et al., 2009; Todgham). и Хофманн, 2009 г., О'Доннелл и др., 2010 г., Спайсер и др., 2011 г., Штумпп и др., 2011 г., Песпени и др., 2013b).Таблица 3
Функциональные классы белков с повышенной представленностью SNP коррелируют с локальными условиями pH ( P < 0,01)
| Функциональная категория . | Идентификатор GO . | № SNP . | № Гены . | P -значение . |
|---|---|---|---|---|
| custom list | 884 | 97 | 0. 001 | |
| Cell–cell adhesion | GO:0016337 | 695 | 63 | 0.001 |
| Proton transport | GO:0015992 | 364 | 45 | 0,002 |
| Окисление липидов | GO: 0034440 | 425 | 37 | 0,003 | 0,003 | 444444444499GO:0006820 | 499 | 60 | 0.003 |
| Fatty acid catabolic process | GO:0009062 | 404 | 35 | 0.008 |
| Homophilic cell adhesion | GO:0007156 | 266 | 30 | 0.008 |
| Response to amine stimulus | GO:0014075 | 182 | 19 | 0.009 |
| Общее количество уникальные гены | 299 |
| Функциональная категория
. | Идентификатор GO . | № SNP . | № Гены . | P -значение . |
|---|---|---|---|---|
| Транспорт протонов, связанный с энергией, против электрохимического градиента | GO:0015988 | 285 | 32 | 0.000 |
| Biomineralization | custom list | 884 | 97 | 0.001 |
| Cell–cell adhesion | GO:0016337 | 695 | 63 | 0.001 |
| Proton transport | GO:0015992 | 364 | 45 | 0.002 |
| Lipid oxidation | GO:0034440 | 425 | 37 | 0.003 |
| Anion transport | GO:0006820 | 499 | 60 | 0.003 |
| Fatty acid catabolic process | GO:0009062 | 404 | 35 | 0,008 |
| гомофильные клеточные адгезии | GO: 0007156 | 266 | 30 | 0,008 |
| Реакция на Amine Strimul Strimul | ||||
| . 0331 | GO:0014075 | 182 | 19 | 0,009 |
| Всего № уникальные гены | 299 |
Открыть в новой вкладке
Таблица 3
Функциональные классы белков с повышенной представленностью SNP, коррелирующих с локальными условиями pH ( P < 0,01)
| Функциональная категория . | Идентификатор GO . | № SNP . | № Гены . | P -значение . |
|---|---|---|---|---|
| Energy coupled proton transport, against electrochemical gradient | GO:0015988 | 285 | 32 | 0.000 |
| Biomineralization | custom list | 884 | 97 | 0.001 |
| Cell–cell adhesion | GO:0016337 | 695 | 63 | 0. 001 |
| Proton transport | GO:0015992 | 364 | 45 | 0.002 |
| Lipid oxidation | GO :0034440 | 425 | 37 | 0.003 |
| Anion transport | GO:0006820 | 499 | 60 | 0.003 |
| Fatty acid catabolic process | GO:0009062 | 404 | 35 | 0.008 |
| Homophilic cell adhesion | GO:0007156 | 266 | 30 | 0.008 |
| Response to amine stimulus | GO:0014075 | 182 | 19 | 0,009 |
| Всего № уникальные гены | 299 |
| Функциональная категория . | Идентификатор GO . | № SNP
. | № Гены . | P -значение . |
|---|---|---|---|---|
| custom list | 884 | 97 | 0.001 | |
| Cell–cell adhesion | GO:0016337 | 695 | 63 | 0.001 |
| Proton transport | GO:0015992 | 364 | 45 | 0,002 |
| Окисление липидов | GO: 0034440 | 425 | 37 | 0,003 | 0,003 | 444444444499GO:0006820 | 499 | 60 | 0.003 |
| Fatty acid catabolic process | GO:0009062 | 404 | 35 | 0.008 |
| Homophilic cell adhesion | GO:0007156 | 266 | 30 | 0.008 |
| Response to amine stimulus | GO:0014075 | 182 | 19 | 0. 009 |
| Общее количество уникальные гены | 299 |
Открыть в новой вкладке
Корреляции SNP с локальным pH и функциональным обогащением могут не выдержать многократную коррекцию тестов из-за высокого уровня потока генов и высокой степени пространственной и временной неоднородности окружающей среды, что ослабляет силу этих признаков естественного отбора на уровне популяции. Однако закономерности сильнее, чем ожидалось, случайно, и поразительно, что гены и функциональные классы белков, наиболее коррелирующие с локальным рН, — это те, которые, по прогнозам, играют роль в росте и выживании при низком рН.
Эти гены также могут быть оценены как биомаркеры для прогнозирования чувствительности или способности к устойчивости определенных групп населения.Рис. 4
Открыть в новой вкладкеСкачать слайд
Гены-кандидаты в реакции на закисление коррелируют с локальным pH и демонстрируют сильную реакцию на экспериментальное закисление в адаптированных к высокому pH популяциях. Частота аллелей представлена в популяции, упорядоченной по рангу времени, проведенного при pH <7,8, обычно с севера на юг, но широту и долготу см. в Таблице 1. Этими местами были Фогарти-Крик, Орегон (ФК), Строберри-Хилл, Орегон (ST), и в Калифорнии в Морском заповеднике Бодега (BD), Государственном парке Ван Дамм (VD), Алегрия, Санта-Барбара (AL) и Террас-Пойнт. Санта-Крус (ТП). Кружки представляют частоты аллелей в природе, оцененные по личинкам, отобранным в день 1 в условиях CO 9 окружающей среды.0181 2 ; ромбы представляют частоты аллелей после экспериментального подкисления, день 7 при повышенном уровне CO 2 . Значения R и P получены из корреляции между частотами аллелей в дикой природе (день 1, окружающий CO 2 , синие точки) и ранговым временем, проведенным при pH <7,8.
Рис. 5
Открыть в новой вкладкеСкачать слайд
Генетические изменения в ответ на экспериментальное закисление максимальны в популяциях с более высоким pH в дикой природе ( t = 4,03, P < 0,0001) у предположительно адаптированных к pH SNP ( n = 318, P < 0,01). Сплошное распределение показывает изменения частоты аллелей в ответ на экспериментальное подкисление в популяциях с низким pH в дикой природе (FC и ST), тогда как штриховое распределение показывает изменения частоты аллелей в ответ на экспериментальное подкисление в популяциях с высоким pH в дикой природе. (АЛ и ТП). Площадь под кривой функции плотности представляет вероятность наблюдения значения x с учетом диапазона значений x (рассчитано с использованием функции «плотность» в R). Ковровые графики под графиком показывают фактические точки данных в виде делений. Генетические изменения между дикой средой (день 1, окружающая среда CO 2 ) и обработкой подкислением (день 7, повышенный уровень CO 2 ) увеличивают частоту аллелей, предположительно адаптированных к низкому pH.
Наши результаты выявили несколько открытых вопросов, касающихся потенциальной устойчивости видов к изменяющимся климатическим условиям. К ним относятся: (1) Какова цена такой быстрой эволюции и последующей утраты генетического разнообразия? (2) Поскольку адаптация требует генетической изменчивости, обладают ли организмы способностью реагировать на одновременные воздействия окружающей среды, такие как изменения pH, температуры, хищников, болезней и наличия пищи? (3) Какие характеристики на уровне вида, такие как размер популяции, уровни генетической изменчивости, поток генов и степень пространственной и временной неоднородности окружающей среды, испытанной в ходе эволюционной истории вида, предсказывают их устойчивость или уязвимость к изменяющимся климатическим условиям?
Наши данные обнадеживают, но для пурпурных морских ежей картина остается неполной. Что касается пурпурных морских ежей и других видов, необходимо больше знать о способности нескольких естественных популяций реагировать на одновременные факторы стресса окружающей среды. Будущие изменения окружающей среды могут быть синергетическими, а не аддитивными, и экстремальные значения на обоих концах температурного и рН-спектра могут быть более распространенными. Например, за последние 50 лет интенсивность апвеллинга увеличивалась, что периодически приводило к более экстремально низким pH и низким температурам, в то время как среднегодовые температуры неуклонно росли (Бакун 19).90; Снайдер и др. 2003 г.; Айлс и др. 2012). Будущие взаимодействия климата и экосистемы будут сложными и неопределенными, а последствия этих взаимодействий могут быть драматичными (Бакун, 1990 г.; Хельмут и др., 2010 г.). Наконец, мы не знаем, как закономерности, которые мы наблюдаем у этого вида, перенесутся на другие виды. Эволюция в изменчивой во времени и пространстве среде может породить генетическое разнообразие, необходимое для эволюционной реакции на изменяющиеся климатические условия, хотя более глубокое понимание способности популяций и видов к быстрым эволюционным реакциям необходимо для прогнозирования последствий климатических изменений и управления ими. .
«> Список литературы
Ashburner
M
,
,
Blake
J
,
Botstein
D
,
D
,
. 0004
Cherry
JM
,
Davis
AP
,
Dolinski
K
,
Dwight
SS
,
Eppig
JT
, et al.
Генная онтология: инструмент для унификации биологии
25
(стр.
25
—
9
)
Bairoch
A
,
Bougueleret
L
,
Altairac
S
,
Amendolia
V
,
Auchincloss
A
,
Argoud-Puy
G
,
Axelsen
K
,
Baratin
D
,
Blatter
MC
,
MC
,
0074 ,
Boeckmann
B
, и др.
Универсальный белковый ресурс (UniProt) 2009
,
Рез. нуклеиновых кислот
,
2009
, vol.
37
(стр.
D169
—
74
)
Бакун
А
4 9.
Глобальное изменение климата и усиление прибрежного апвеллинга океана
,
Наука
,
1990
, том.
247
(стр.
198
—
201
)
Bakun
A
,
Parrish
RH
.
Турбулентность, перенос и пелагическая рыба в системах течений Калифорнии и Перу
23
(стр.
99
—
112
)
Benjamini
Y
,
Hochberg
Y
4 .
Контроль частоты ложных обнаружений: практичный и эффективный подход к множественному тестированию
51
(стр.
289
—
300
)
Biermann
CH
,
Кессинг
BD
,
Палумби
SR
.
Филогения и развитие морских модельных видов: стронгилоцентроидные морские ежи
5
(стр.
360
—
71
)
Byrne
M
,
Selvakumaraswamy
P
,
. 0004
,
,
Вулси
E
,
Нгуен
H
.
Развитие морского ежа в очаге глобальных изменений, возможность миграции термоустойчивых пропагул к югу
58
(стр.
712
—
9
)
De Wit
P
,
Pespeni
MH
,
Ladner
JT
,
Barshis
DJ
,
Seneca
F
,
Jaris
H
,
Теркильдсен
NO
,
Морикава
M
,
Палумби
SR
4 SR
4 .
Простое руководство для дураков по популяционной геномике с помощью RNA-Seq: введение в анализ данных высокопроизводительного секвенирования
,
Мол Экол Ресурс
,
2012
, том.
12
(pg.
1058
—
67
)
Doney
SC
,
Fabry
VJ
,
Feely
RA
,
Kleypas
JA
.
Подкисление океана: другой CO 2 проблема
,
Ann Rev Mar Sci
,
2009
, vol.
1
(pg.
169
—
92
)
Edmands
S
,
Moberg
PE
,
Burton
RS
.
Аллозимная и митохондриальная ДНК, свидетельствующие о подразделении популяции пурпурного морского ежа Strongylocentrotus purpuratus
,
Mar Biol
,
1996
, vol.
126
(стр.
443
—
50
)
Endler
JA
4 .
Географические вариации, видообразование и клины
,
1977
Принстон (NJ)
Princeton University Press
Estes
JA
,
Steinberg
PD
.
Хищничество, травоядность и эволюция водорослей
,
Палеобиология
,
1988
, том.
14
(стр.
19
—
36
)
Evans
TG
,
,
9777777. Транскриптомные реакции на закисление океана у личинок морских ежей в среде с естественным переменным рН
,
Мол Экол
,
2013
, том.
22
(pg.
1609
—
25
)
Feely
RA
,
Sabine
CL
,
Hernandez-Ayon
JM
,
Янсон
Д
,
Хейлз
Б
.
Доказательства подъема коррозионно-активных «подкисленных» вод на континентальный шельф
,
Наука
,
2008
, том.
320
стр.
1490
Findlay
HS
,
Calosi
P
,
Crawfurd
K
.
Детерминанты ПОС: реакция ВОУ кокколитофориды Emiliania huxleyi при будущих сценариях закисления океана
,
Limnol Oceanogr
,
2011
, том.
56
стр.
1168
,
Dworjanyn
SA
,
POORE
AGB
,
Byrne
M
.
Адаптивная способность среды обитания модифицировать морского ежа Centrostephanus rodgersii к потеплению и закислению океана: характеристики ранних эмбрионов
,
PLoS One
,
2012
, том.
7
стр.
E42497
Garfield
D
,
Haygood
R
,
Nielsen
WJ
,
WRAY
GA
4444444444444444444444444444444444444444. 4004444444444444444. Популяционная генетика цис-регуляторных последовательностей, действующих во время эмбрионального развития морского ежа Strongylocentrotus purpuratus
,
Эвол Дев
,
2012
, том.
14
(стр.
152
—
67
)
Gilchrist
GW
,
Huey
RB
.
Пластическая и генетическая изменчивость нагрузки на крыло в зависимости от температуры внутри и между параллельными клинами в Drosophila subobscura
,
Integr Comp Biol
,
2004
, том.
44
(pg.
461
—
70
)
Harley
CDG
,
Randall Hughes
A
,
Hultgren
KM
,
Шахтер
BG
,
Sorte
CJ
,
Торнбер
CS
4
4 Родригес0002 LF
,
Томанек
L
,
Williams
SL
.
Влияние изменения климата на прибрежные морские системы
,
Ecol Lett
,
2006
, vol.
9
(стр.
228
—
41
)
Helmuth
B
,
Broitman
Br
,
0004
Yamane
L
,
Gilman
SE
,
,
Mislan
KA
,
Denny
MW
444444444444444444444444444444444444444444444444444444444444.
Климатология организма: анализ изменчивости окружающей среды в масштабах, соответствующих физиологическому стрессу
213
(стр.
995
—
1003
)
Hermisson
J
,
Pennings
P
.
Мягкие зачистки: молекулярная популяционная генетика адаптации от постоянной генетической изменчивости
169
стр.
2335
Хочачка
ПВ
,
Сомеро
ГН
.
Biochemical adaptation: mechanism and process in physiological evolution
,
2002
New York (NY)
Oxford University Press
Hoegh-Guldberg
O
,
Mumby
P
,
Hooten
A
,
Steneck
RS
,
Greenfield
P
,
GOMEZ
E
,
Gomez
E
,
Harvell
CD
,
Продажа
PF
,
Edwards
AJ
,
Caldeira
K
, at.
Коралловые рифы в условиях быстрого изменения климата и закисления океана
318
стр.
1737
Хоффманн
АА
,
Sgrò
CM
.
Изменение климата и эволюционная адаптация
,
Природа
,
2011
, vol.
470
(pg.
479
—
85
)
Hofmann
GE
,
Smith
JE
,
Johnson
KS
,
,
Левин
LA
,
Micheli
F
,
Paytan
A
,
Price
NN
,
Peterson
B
,
Takeshita
Y
, и другие.
Высокочастотная динамика рН океана: сравнение нескольких экосистем
6
стр.
e28983
Iles
AC
,
Gouhier
TC
,
Menge
BA
,
Stewart
JS
,
Haupt
AJ
,
Линч
МС
.
Тенденции, определяемые климатом, и экологические последствия апвеллинга масштаба события в Калифорнийской системе течений
,
Glob Change Biol
,
2012
, том.
18
(стр.
783
—
96
)
Jacobs
DK
,
9000 244
4494
449н. Гены, разнообразие и геологические процессы на побережье Тихого океана
,
Annu Rev Earth Planet Sci
,
2004
, vol.
32
(pg.
601
—
52
)
Kelly
MW
,
Padilla‐Gamiño
JL
,
Hofmann
GE
.
Естественная изменчивость и способность адаптироваться к закислению океана у краеугольного морского ежа.
19
(стр.
2536
—
46
)
, 7
A
4
4 Эволюция на двух уровнях у человека и шимпанзе
188
(стр.
107
—
16
)
Lande
R
,
Shannon
S
,
0004
.
Роль генетической изменчивости в адаптации и сохранении популяции в меняющейся среде
50
(pg.
434
—
7
)
Langer
G
,
Nehrke
G
,
Probert
I
,
,
Зивери
Р
.
Штамм-специфические реакции Emiliania huxleyi на изменение химического состава карбонатов морской воды
6
(стр.
2637
—
46
)
,
Braynen
W
,
Keshav
K
44444444,.444444444474, .
ErmineJ: инструмент для функционального анализа наборов данных об экспрессии генов
6
стр.
269
.
Молекулярные филогении и времена дивергенции видов морских ежей Strongylocentrotidae, Echinoida
,
Mol Biol Evol
,
2003
, vol.
20
(стр.
1211
—
21
)
Levine
MT
,
Eckert
ML
,
Betun
DJ
.
Полногеномная пластичность экспрессии в тропиках и умеренных широтах Drosophila melanogaster популяций из Восточной Австралии
28
(стр.
249
—
56
)
,
Дурбин
R
.
Быстрое и точное выравнивание коротких прочтений с помощью преобразования Берроуза-Уилера
25
(стр.
1754
—
60
)
Mann
KH
,
Lazier
JRN
,
0004
.
Dynamics of marine ecosystems: biological-physical interactions in the oceans
,
2006
Nova Scotia, Canada
Wiley-Blackwell
Martz
TR
,
Connery
JG
,
Джонсон
КС
.
Испытание Honeywell Durafet® на определение pH морской воды
,
Limnol Oceanogr Methods
,
2010
, том.
8
(pg.
172
—
84
)
McKenna
A
,
Hanna
M
,
Banks
E
,
Sivachenko
A
,
Cibulskis
K
,
Kernytsky
A
,
Garimella
K
,
Альтшулер
D
,
Габриэль
S
,
, и др.
Набор инструментов для анализа генома: платформа MapReduce для анализа данных секвенирования ДНК нового поколения
20
(стр.
1297
—
303
)
Мельцнер
,
0004
Gutowska
M
,
Langenbuch
M
,
Dupont
S
,
Lucassen
M
,
Thorndyke
MC
,
Bleich
M
,
Pörtner
H-O
.
Физиологическая основа высокой толерантности к СО2 у морских экзотермических животных: предварительная адаптация посредством образа жизни и онтогенеза?
,
Биогеонауки
,
2009
, том.
6
(стр.
4693
—
738
)
Менге
BA
4 .
Регулирование сообществ сверху вниз и снизу вверх в морских каменистых литоральных местообитаниях
250
(стр.
257
—
89
)
Moss
RH
,
Edmonds
JA
,
Hibbard
KA
,
Manning
MR
,
Rose
SK
,
Van Vuuren
DP
,
Carter
TR
,
Emori
S
,
Kainuma
M
,
M
,
M
,
M
,
. 0004
,
и др.
Сценарии следующего поколения для исследования и оценки изменения климата
463
(pg.
747
—
56
)
O’Donnell
MJ
,
Todgham
AE
,
Sewell
MA
.
Подкисление океана изменяет скелетогенез и экспрессию генов у личинок морских ежей
,
Mar Ecol Prog Ser
,
2010
, vol.
398
(pg.
157
—
71
)
Oksanen
J
,
Kindt
R
,
Legendre
P
,
O ‘Хара
РБ
,
Симпсон
GL
,
Стивенс
MHH
. ,
2008
Палумби
SR
,
Уилсон
AC
.
Разнообразие митохондриальной ДНК морских ежей Strongylocentrotus purpuratus и S
,
droebachiensis . Эволюция
,
1990
, том.
44
(стр.
403
—
15
)
Паркер
L
,
Росс
PM
,
О’Коннор
WA
.
Популяции сиднейской каменной устрицы, Saccostrea glomerata , изменяются в зависимости от закисления океана
158
(стр.
689
—
97
)
Песпени
MH
,
Барни
BT
,
Палумби
SR
.
Различия в регуляции генов роста и биоминерализации, выявленные в результате многолетней обычной садовой акклиматизации и экспериментальной геномики у пурпурного морского ежа
67
(стр.
1901
—
14
)
Pespeni
MH
,
Garfield
DA
,
Manier
Mk
,
Palumbi
SR
.
Полногеномные полиморфизмы обнаруживают неожиданные цели естественного отбора
279
(стр.
1412
—
20
)
Песни
MH
,
Oliver
TA
,
Manier
MK
,
Palumbi
SR
.
Анализ тайлинга сайтов рестрикции: точное обнаружение и количественное генотипирование полиморфизмов всего генома с использованием массивов нуклеотидов
11
стр.
Р44
Песпени
MH
,
Палумби
SR
.
Сигналы отбора в аномальных локусах у широко расселенных видов в градиенте окружающей среды
22
(стр.
3580
—
97
)
Песни
MH
,
Сэнфорд
0004 E
,
Gaylord
B
,
Hill
TM
,
Hosfelt
JD
,
Jaris
HK
,
LaVigne
M
,
Lenz
EA
,
Russell
AD
,
Young
MK
, 9007 и др. Эволюционные изменения во время экспериментального закисления океана
,
Proc Natl Acad Sci USA
,
2013b
, vol.
110
(pg.
6937
—
42
)
Pistevos
JCA
,
Calosi
P
,
Widdicombe
S
,
Bishop
JDD
.
Будут ли различия среди генетических особей влиять на реакцию видов на глобальное изменение климата?
,
Ойкос
,
2011
, том.
120
(стр.
675
—
89
)
Pörtner
H-O 90.0 90.
Экосистемные эффекты закисления океана во время его потепления: взгляд физиолога
373
(стр.
203
—
17
)
Pörtner
HO
,
Langenbuch
M
,
Michaelidis
B
.
Синергетическое воздействие экстремальных температур, гипоксии и повышения концентрации CO 2 на морских животных: от истории Земли к глобальным изменениям
110
стр.
C09S10
Проссер
CL
.
Адаптационная биология: молекулы для организмов
,
1986
Нью -Йорк (Нью -Йорк)
Wiley
R Основная команда разработки
R: Язы
Австрия: R Foundation for Statistical Computing
Ridgwell
A
,
Schmidt
D
,
Turley
C
,
Brownlee
C
,
Maldonado
MT
,
Tortell
P
,
Young
JR
.
От лабораторных манипуляций к моделям системы Земля: масштабирование последствий кальцификации подкисления океана
,
Биогеонауки
,
2009
, том.
6
(стр.
2611
—
23
)
Rogers-Bennett
7 L4
2 Лоуренс
ДМ
.
Экология Strongylocentrotus franciscanus и Strongylocentrotus purpuratus
,
Съедобные морские урчины: биология и экология
,
2007
Amsterm
,
2007
Amsterm9
004
(стр.
393
—
425
)
Санфорд
E
,
Kelly
M
.
Местная адаптация у морских беспозвоночных
3
(стр.
509
—
35
)
Selkoe
K
,
Watson
J
,
White
C
,
Horin
TB
,
Iacchei
M
,
Mitarai
S
,
Siegel
DA
,
Гейнс
SD
,
Тунен
RJ
.
Преодоление хаоса генетической мозаичности: генетика морских ландшафтов выявляет экологические и океанографические факторы, определяющие генетические закономерности у трех видов рифов умеренного пояса
,
Мол Экол
,
2010
, том.
19
стр.
3708
Слаткин
М
.
Поток генов и географическая структура природных популяций
236
(стр.
787
—
92
)
Smith
PE
,
Эппли
RW
.
Первичная продукция и популяция анчоуса в Южно-Калифорнийском заливе: сравнение временных рядов
27
(стр.
1
—
17
)
Snyder
MA
,
Sloan
LC
,
.0002 Diffenbaugh
NS
,
Bell
JL
.
Будущие изменения климата и апвеллинг в Калифорнийском течении
,
Geophys Res Lett
,
2003
, vol.
30
Spicer
J
,
Widdicombe
S
,
Needham
H
,
Berge
J
H
,
H
,
.0004
.
Влияние CO 2 -Ацидифицированная морская вода внеклеточной кислоты -базы северного морского ежа Стронглоцентрот Dröebachiensis
,
J exp Mar Biol ECOL
,
1114
9000 2.
407
(стр.
19
—
25
)
Strathmann
MF4 MF.
В кн.: Размножение и развитие морских беспозвоночных северного побережья Тихого океана: данные и методы изучения яиц, зародышей и личинок
,
1987
Сиэтл (WA)
Университет Вашингтонской прессы
(стр.
511
—
34
)
Strathmann
R 9000 40074.
Продолжительность пелагического периода у иглокожих с питающимися личинками из северо-восточной части Тихого океана
34
(стр.
23
—
8
)
Stumpp
M
,
,
Melzner
F
,
Thorndyke
M
,
Dupont
S
.
CO 2 Индуцированное окисление морской воды влияет на развитие личинок морского ежа I: повышенная скорость метаболизма снижает возможности роста и вызывает задержку развития
,
Comp Biochem Physiol A Mol Integr Physiol
,
2011
, том.
160
(pg.
331
—
40
)
Sunday
JM
,
Crim
RN
,
Harley
CDG
,
Hart
МВт
.
Количественная оценка скорости эволюционной адаптации в ответ на закисление океана
,
PLoS One
,
2011
, том.
6
стр.
E22881
Swindell
W
,
Huebner
M
,
Weber
A
.
Паттерны пластической и адаптивной экспрессии генов, связанные с температурным стрессом у Arabidopsis thaliana
,
Наследственность
,
2007
, vol.
99
(pg.
143
—
50
)
Szászi
K
,
Grinstein
S
,
Orlowski
J
,
Kapus
А
.
Регуляция эпителиальной изоформы обменника Na+/H+ цитоскелетом
10
(стр.
265
—
72
)
Todgham
AE
,
Hofmann
GE
.
Транскриптомный ответ личинок морского ежа Строильный заводрский пурпурат на CO 2 -Осгивание морской воды
,
J Exp Biol
,
2009
, кет.
212
(стр.
2579
—
94
)
Vermeij
GJ
.
Географическое ограничение как руководство к причинам вымирания: случай холодных северных океанов в неогене
15
(стр.
335
—
56
)
Whitehead
A
,
Crawford
ДЛ
.
Изменчивость внутри и между видами в экспрессии генов: сырье для эволюции
15
(pg.
1197
—
211
)
Zaytsev
O
,
Cervantes-Duarte
R
,
Montante
O
,
Гальегос-Гарсия
А
.
Прибрежный апвеллинг на тихоокеанском шельфе полуострова Нижняя Калифорния
59
(стр.
489
—
502
)
Приложение 1
Методы
Сборник из Урчинса
THIRTO 1) и отправлен в морскую лабораторию Bodega. Для каждой популяции ежей засевали 0,5 М KCl в морской воде и смешивали гаметы 10 самок и 10 самцов для оплодотворения. Оплодотворенные эмбрионы из каждой популяции были разделены на две группы, и группы были помещены в два отдельных сосуда, один из которых поддерживался при температуре окружающей среды p CO 2 уровни (400 мкатм, современный глобальный средний атмосферный CO 2 ), а другой — повышенные уровни p CO 2 (900 мкатм, умеренный прогноз «интенсивного использования ископаемого топлива» на 2100 год) (Мосс и др., 2010). Культуры содержались с плотностью 0,66 личинок на миллилитр, что соответствует экологически значимой плотности (Strathmann, 1987). Сосуды с культурами держали в столах с морской водой при температуре 14°C. Морскую воду в сосудах постоянно перемешивали вибрирующими лопастями и каждые 2 дня 90% морской воды заменяли, удаляя воду обратной фильтрацией через сетку и заменяя отфильтрованной морской водой, предварительно уравновешенной до желаемого уровня p CO 2 . Начиная с 3-х суток после оплодотворения после каждого водообмена личинок кормили смесью водорослей ( Rhodomonas sp. и Dunaliella sp.) ad libitum . См. Песпени и др. (2013b) для получения полной информации о методах культивирования и химическом составе воды.
Популяционная геномика
Подводя итоги предыдущего исследования, можно сказать, что для каждой популяции мы отобрали примерно 1000 живых личинок на 1-й и 7-й день после оплодотворения для каждой обработки p CO 2 . Мы экстрагировали общую РНК из каждого пула личинок с помощью TRIzol и подготовили библиотеки кДНК для секвенирования с использованием набора TruSeq в соответствии с протоколом производителя (Illumina, Сан-Диего, Калифорния). Каждый образец был секвенирован на одной дорожке Illumina HiSeq, что дало примерно 80 миллионов прочтений по 50 пар оснований на образец (Microarray and Genomics Core Facility, Университет Юты, Солт-Лейк-Сити, Юта). Необработанные данные последовательности были обрезаны по качеству, а затем по длине с использованием программ набора инструментов FASTX (Gordon 2011) с минимальной оценкой качества по шкале Phred 30 и минимальной длиной 30 п.н. Мы сопоставили прочтения каждого образца со всеми предсказанными генами пурпурного морского ежа (загружены с www.spbase.org), используя программу Burrows-Wheeler Aligner (BWA) (Li and Durbin 2009).) и с использованием параметров по умолчанию, кроме n = 0,001, чтобы учесть высокую генетическую изменчивость пурпурных морских ежей (Консорциум по секвенированию генома морских ежей и др., 2006; Песпени и Палумби, 2013). SNP во всех образцах были обнаружены с использованием программ набора инструментов для анализа генома (GATK) (McKenna et al., 2010) и самых строгих критериев разработчика (http://www.broadinstitute.org/gatk/guide/). Оценки частоты аллелей для каждого образца личиночного пула были извлечены из полученных файлов .vcf путем деления количества чтений качества, сопоставленных с эталонным аллелем, на общее количество сопоставленных чтений качества в этой позиции с использованием пользовательского скрипта Python. (Доступен по запросу). Список этапов обработки данных, которые можно настроить, можно найти на сайте http://sfg.stanford.edu (De Wit et al. 2012).
Оценки частот аллелей у личинок, отобранных в день 1 в условиях окружающей среды CO 2 , были взяты для представления частот аллелей, присутствующих в диких популяциях. Оценки личинок, отобранных на 7-й день после культивирования при повышенном уровне CO 2 , были взяты для представления частот аллелей после лабораторного культивирования при высоком уровне CO 2 . Различия в частоте аллелей между этими двумя образцами «день × лечение» были взяты для представления изменений в частоте аллелей из-за смертности личинок. Альтернативными объяснениями различий в частоте аллелей могут быть (1) различия в размере личинок и, следовательно, в обилии транскриптов среди образцов (однако мы измерили минимальные различия в размерах между обработками, поэтому размер личинок вряд ли повлияет на оценки) или (2 ) различия в аллель-специфической экспрессии, но мы не нашли доказательств этого в данных (см. Pespeni et al. (2013b) для анализа и обсуждения).
Сбор данных о рН окружающей среды
Мы провели высокочастотные измерения локального рН с использованием автономного датчика рН, установленного на каждом участке исследования (Ф. Чан и др., в процессе подготовки). Датчик pH был установлен в приливных участках, где собирались взрослые морские ежи, чтобы наиболее точно регистрировать pH, с которым сталкиваются ежи в дикой природе (за исключением Алегрии, Калифорния, 34,47 ° с.ш., где записи с ближайшего доступного датчика в Ломпоке, Калифорния , 34,72° с.ш.). Датчик pH представляет собой модифицированную версию датчика SeaFET (Martz et al. 2010), в котором используется pH-зонд Honeywell DuraFET с ионно-чувствительным полевым транзистором, встроенный регистратор данных и блок питания (Evans et al. 2013). Измерения проводились каждые 10 минут в течение сезона апвеллинга с апреля по сентябрь 2011 года в керне. Количество времени, проведенное ниже pH 7,8, было рассчитано для каждого участка, чтобы отразить серьезность низкого pH, наблюдаемого на каждом участке. Участки были ранжированы с 1 по 6 в зависимости от времени, проведенного при pH ниже 7,8, при этом 1 провел больше всего времени при низком pH (таблица 1).
Тест на корреляцию между частотой аллеля и локальным рН
Для каждого SNP мы проверили корреляцию между частотой аллеля и ранговым локальным рН в шести местах исследования. Мы использовали частичные тесты Mantel, реализованные в среде программирования R (R Development Core Team 2009), с использованием функции «mantel.partial» в веганской библиотеке (Oksanen et al. 2008), контролирующей географическое расстояние. P -значения определялись путем случайной перестановки данных и пересчета корреляции 1000 раз. Перед проверкой корреляции мы стандартизировали дисперсию для каждой переменной, разделив ее на среднее значение и вычитая одно стандартное отклонение в соответствии с процедурой Selkoe et al. (2010).
Тест на взаимосвязь между функцией гена и корреляцией с локальным pH
Мы проверили неслучайную концентрацию SNP с высокой корреляцией с локальными условиями pH в определенных функциональных классах генов, подход, называемый анализом функционального обогащения. Мы охарактеризовали каждый ген, используя идентификаторы UniProt (Bairoch et al. 2009), затем категории биологических процессов Gene Ontology (GO) (Ashburner et al. 2000). Мы также включили список генов биоминерализации, генов, важных для роста скелета морских ежей (как в Pespeni et al. 2013a, 2013b). Чтобы уменьшить шум и сосредоточиться на более интересных паттернах генома, мы ограничили анализ функциональными классами, содержащими от 10 до 100 членов генов, исключая классы, которые были слишком узкими или широкими, соответственно. Было 1307 функциональных категорий, которые содержали от 10 до 100 генов-членов. Для каждой функциональной категории мы проверили корреляцию между принадлежностью к этой категории и оценкой гена, используя повторную выборку оценки гена, реализованную в ErmineJ (Lee et al. 2005). Для оценок генов мы использовали 9 логарифмических преобразований.0014 P — значение для каждого SNP, рассчитанное в частичном тесте Мантеля для корреляции между частотой аллеля и ранговым pH, описанным выше. Когда было несколько SNP на ген, мы использовали средний балл. Статистическая значимость определялась по 10 000 перестановок, и скорректированные значения P рассчитывались с использованием подхода Бенджамини-Хохберга (Бенджамини и Хохберг, 1995). Чтобы определить, были ли определенные функциональные категории чрезмерно представлены среди тех, у кого P <0,01, мы сравнили наблюдаемые частоты с ожидаемыми частотами выше и ниже порогового значения 9.0014 P — значение с использованием точного критерия Фишера.
Идентификация лучших генов-кандидатов и сигналов селекции
Мы идентифицировали главные гены-кандидаты для реакции на закисление путем изучения 318 SNP с корреляцией с локальным pH ( P < 0,01). Эти SNP продемонстрировали общие клинальные паттерны в отношении рангового времени, проведенного при pH <7,8 для каждого сайта. Мы сравнили частоты аллелей, рассчитанные по клинальным аллелям в дикой природе (день 1, 400 мкАТм), с частотами аллелей через 7 дней при низком pH (день 7; 9 дней). 00 мкатм). Предполагая, что эти аллели адаптированы к низкому рН, мы искали минимальные изменения частоты аллелей при экспериментальном закислении в популяциях, предположительно адаптированных к низкому рН (FC и ST), и более значительные изменения в популяциях с высоким рН (AL и TP). ). Если бы все популяции отреагировали на экспериментальное подкисление к 7-му дню, можно было бы ожидать низкой изменчивости частот аллелей среди популяций на 7-й день для личинок при 900 мкатм. Чтобы определить лучших кандидатов на реакцию на подкисление, мы сделали эти расчеты, как описано, и соответственно ранжировали локусы. Мы также визуально изучили паттерны во всех 318 локусах, написав сценарий для циклического просмотра и построения всех 318 локусов. На основе этих критериев были отобраны семь лучших кандидатов, хотя все 318 локусов с частотами аллелей для каждой популяции можно найти в дополнительной таблице S1.
Если бы экспериментальная селекция была нацелена на те же аллели, которые были адаптированы к местным условиям pH в дикой природе, мы могли бы ожидать, что степень генетических изменений в ответ на экспериментальное закисление (низкий pH, высокий уровень CO 2 ) будет наибольшей в популяциях, которые обычно имеют высокий pH в природе и более низкий в популяциях, которые обычно имеют низкий pH в природе. Чтобы проверить это, мы рассчитали среднюю разницу в частоте аллелей между личинками на 1-й день (400 мкатм) и личинками на 7-й день (900 мкатм) в наборах из двух популяций с высшим и низшим ранговыми условиями рН (FC и ST, AL и TP соответственно). Мы сравнили два распределения ответов, используя критерий Колмогорова-Смирнова (K-S) (функция ks.test в R), и сравнили эти средние ответы в условиях низкого pH, используя t -критерий, предсказывающий, что популяции с высоким рангом проявят наибольший отклик. (функция t.test в R).
Примечания автора
Из симпозиума «Понимание фенотипов первого порядка: транскриптомика для новых моделей», представленного на ежегодном собрании Общества интегративной и сравнительной биологии 3–7 января 2013 г. в Сан-Франциско, Калифорния.
© Автор, 2013 г. Опубликовано Oxford University Press от имени Общества интегративной и сравнительной биологии. Все права защищены. Для получения разрешений обращайтесь по электронной почте: journals. [email protected].
© Автор, 2013 г. Опубликовано Oxford University Press от имени Общества интегративной и сравнительной биологии. Все права защищены. Для получения разрешений обращайтесь по электронной почте: [email protected].
Раздел выпуска:
Понимание фенотипов первого порядка: транскриптомика для новых моделей
Скачать все слайды
Дополнительные данные
Дополнительные данные
Дополнительные данные — zip-файл
Реклама
Цитаты
Альтметрика
Дополнительная информация о метриках
Оповещения по электронной почте
Оповещение об активности статьи
Предварительные уведомления о статьях
Оповещение о новой проблеме
Получайте эксклюзивные предложения и обновления от Oxford Academic
Ссылки на статьи по телефону
Последний
Самые читаемые
Самые цитируемые
Экоиммунология: что нетрадиционные организмы говорят нам через два десятилетия
Биотические взаимодействия и будущее рыб на коралловых рифах: важность подходов, основанных на признаках
Потепление ускоряет начало реакции на молекулярный стресс и увеличивает смертность личинок атлантической трески
Шаг вперед: функциональное разнообразие и новые темы медленной локомоции у позвоночных
Лучшие практики биоинспирированного дизайна: ключевые темы и проблемы
Реклама
Реакция вашего организма на стресс
Обзор
Стресс — обычное явление. Хотя вы не можете удалить из своей жизни все факторы стресса, можно управлять стрессом и поддерживать свое здоровье. Это важно, потому что стресс может вызвать умственную усталость, раздражительность и бессонницу.
Но даже если вы знаете о физических последствиях стресса, вы можете не знать о различных стадиях стресса, известных как общий адаптационный синдром (ОАС). Когда вы понимаете разные стадии стресса и то, как организм реагирует на эти стадии, вам будет легче выявить у себя признаки хронического стресса.
Подробнее: 20 эффектов стресса на организм »
ГАЗ представляет собой трехэтапный процесс, который описывает физиологические изменения, происходящие в организме в условиях стресса. Ганс Селье, врач и исследователь, выдвинул теорию ГАС. Во время эксперимента с лабораторными крысами в Университете Макгилла в Монреале он наблюдал ряд физиологических изменений у крыс после того, как они подверглись стрессовым событиям.
С помощью дополнительных исследований Селье пришел к выводу, что эти изменения были не изолированным случаем, а скорее типичной реакцией на стресс. Селье определил эти стадии как тревогу, сопротивление и истощение. Понимание этих различных реакций и того, как они соотносятся друг с другом, может помочь вам справиться со стрессом.
Подробнее: 10 простых способов снять стресс »
1. Стадия реакции тревоги
Стадия реакции тревоги относится к начальным симптомам, которые испытывает организм при стрессе. Возможно, вы знакомы с реакцией «бей или беги», которая является физиологической реакцией на стресс. Эта естественная реакция подготавливает вас либо к бегству, либо к защите в опасных ситуациях. Ваш сердечный ритм увеличивается, надпочечники выделяют кортизол (гормон стресса), и вы получаете заряд адреналина, который увеличивает энергию. Эта реакция «бей или беги» возникает на стадии реакции тревоги.
2. Стадия сопротивления
После первоначального шока от стрессового события и реакции «бей или беги» тело начинает восстанавливаться. Он высвобождает меньшее количество кортизола, и ваш сердечный ритм и кровяное давление начинают нормализоваться. Хотя ваше тело входит в эту фазу восстановления, некоторое время оно остается в состоянии повышенной готовности. Если вы преодолеваете стресс и ситуация больше не является проблемой, ваше тело продолжает восстанавливаться до тех пор, пока уровень гормонов, частота сердечных сокращений и кровяное давление не достигнут предстрессового состояния.
Некоторые стрессовые ситуации продолжаются длительное время. Если вы не избавитесь от стресса и ваше тело будет оставаться в состоянии повышенной готовности, оно в конечном итоге адаптируется и научится жить с более высоким уровнем стресса. На этом этапе тело претерпевает изменения, о которых вы не подозреваете, пытаясь справиться со стрессом.
Ваше тело продолжает выделять гормон стресса, и ваше кровяное давление остается повышенным. Вы можете думать, что хорошо справляетесь со стрессом, но физическая реакция вашего тела говорит о другом. Если стадия резистентности продолжается слишком долго без пауз, чтобы компенсировать последствия стресса, это может привести к стадии истощения.
К признакам стадии резистентности относятся:
- раздражительность
- разочарование
- плохая концентрация
3. Стадия истощения
Эта стадия является результатом длительного или хронического стресса. Борьба со стрессом в течение длительного времени может истощить ваши физические, эмоциональные и умственные ресурсы до такой степени, что у вашего тела больше не будет сил бороться со стрессом. Вы можете сдаться или почувствовать, что ваше положение безнадежно. К признакам истощения относятся:
- усталость
- выгорание
- депрессия
- тревога
- снижение стрессоустойчивости
Физические эффекты этой стадии также ослабляют вашу иммунную систему и подвергают вас риску заболеваний, связанных со стрессом.
ГАЗ может возникнуть при любом типе стресса. К стрессовым событиям относятся:
- потеря работы
- проблемы со здоровьем
- финансовые проблемы
- распад семьи
- травма
Но, несмотря на то, что стресс неприятен, положительная сторона заключается в том, что GAS улучшает реакцию организма на стрессоры, особенно на стадии тревоги.
Реакция «бей или беги», возникающая на стадии тревоги, предназначена для вашей защиты. Более высокий уровень гормонов на этом этапе приносит вам пользу. Это дает вам больше энергии и улучшает вашу концентрацию, чтобы вы могли сосредоточиться и справиться с ситуацией. Когда стресс кратковременный или кратковременный, стадия тревоги не опасна.
При длительном стрессе это не так. Чем дольше вы справляетесь со стрессом, тем вреднее он наносит вашему здоровью. Вы также не хотите оставаться на стадии сопротивления слишком долго и рискуете войти в стадию истощения. Когда вы находитесь на стадии истощения, продолжительный стресс повышает риск хронического высокого кровяного давления, инсульта, сердечных заболеваний и депрессии. У вас также более высокий риск инфекций и рака из-за более слабой иммунной системы.
Поскольку устранить все факторы стресса невозможно, важно найти способы справиться со стрессом. Знание признаков и стадий стресса может помочь вам предпринять соответствующие шаги, чтобы контролировать уровень стресса и снизить риск осложнений.
Эволюция на двух уровнях у человека и шимпанзе
188
(стр.
107
—
16
)
Lande
R
,
Shannon
S
,
0004
.
Роль генетической изменчивости в адаптации и сохранении популяции в меняющейся среде
50
(pg.
434
—
7
)
Langer
G
,
Nehrke
G
,
Probert
I
,
,
Зивери
Р
.
Штамм-специфические реакции Emiliania huxleyi на изменение химического состава карбонатов морской воды
6
(стр.
2637
—
46
)
,
Braynen
W
,
Keshav
K
44444444,.444444444474,.
ErmineJ: инструмент для функционального анализа наборов данных об экспрессии генов
6
стр.
269
.
Молекулярные филогении и времена дивергенции видов морских ежей Strongylocentrotidae, Echinoida
,
Mol Biol Evol
,
2003
, vol.
20
(стр.
1211
—
21
)
Levine
MT
,
Eckert
ML
,
Betun
DJ
.
Полногеномная пластичность экспрессии в тропиках и умеренных широтах Drosophila melanogaster популяций из Восточной Австралии
28
(стр.
249
—
56
)
,
Дурбин
R
.
Быстрое и точное выравнивание коротких прочтений с помощью преобразования Берроуза-Уилера
25
(стр.
1754
—
60
)
Mann
KH
,
Lazier
JRN
,
0004
.
Dynamics of marine ecosystems: biological-physical interactions in the oceans
,
2006
Nova Scotia, Canada
Wiley-Blackwell
Martz
TR
,
Connery
JG
,
Джонсон
КС
.
Испытание Honeywell Durafet® на определение pH морской воды
,
Limnol Oceanogr Methods
,
2010
, том.
8
(pg.
172
—
84
)
McKenna
A
,
Hanna
M
,
Banks
E
,
Sivachenko
A
,
Cibulskis
K
,
Kernytsky
A
,
Garimella
K
,
Альтшулер
D
,
Габриэль
S
,
, и др.
Набор инструментов для анализа генома: платформа MapReduce для анализа данных секвенирования ДНК нового поколения
20
(стр.
1297
—
303
)
Мельцнер
,
0004
Gutowska
M
,
Langenbuch
M
,
Dupont
S
,
Lucassen
M
,
Thorndyke
MC
,
Bleich
M
,
Pörtner
H-O
.
Физиологическая основа высокой толерантности к СО2 у морских экзотермических животных: предварительная адаптация посредством образа жизни и онтогенеза?
,
Биогеонауки
,
2009
, том.
6
(стр.
4693
—
738
)
Менге
BA
4 .
Регулирование сообществ сверху вниз и снизу вверх в морских каменистых литоральных местообитаниях
250
(стр.
257
—
89
)
Moss
RH
,
Edmonds
JA
,
Hibbard
KA
,
Manning
MR
,
Rose
SK
,
Van Vuuren
DP
,
Carter
TR
,
Emori
S
,
Kainuma
M
,
M
,
M
,
M
,
. 0004
,
и др.
Сценарии следующего поколения для исследования и оценки изменения климата
463
(pg.
747
—
56
)
O’Donnell
MJ
,
Todgham
AE
,
Sewell
MA
.
Подкисление океана изменяет скелетогенез и экспрессию генов у личинок морских ежей
,
Mar Ecol Prog Ser
,
2010
, vol.
398
(pg.
157
—
71
)
Oksanen
J
,
Kindt
R
,
Legendre
P
,
O ‘Хара
РБ
,
Симпсон
GL
,
Стивенс
MHH
. ,
2008
Палумби
SR
,
Уилсон
AC
.
Разнообразие митохондриальной ДНК морских ежей Strongylocentrotus purpuratus и S
,
droebachiensis . Эволюция
,
1990
, том.
44
(стр.
403
—
15
)
Паркер
L
,
Росс
PM
,
О’Коннор
WA
.
Популяции сиднейской каменной устрицы, Saccostrea glomerata , изменяются в зависимости от закисления океана
158
(стр.
689
—
97
)
Песпени
MH
,
Барни
BT
,
Палумби
SR
.
Различия в регуляции генов роста и биоминерализации, выявленные в результате многолетней обычной садовой акклиматизации и экспериментальной геномики у пурпурного морского ежа
67
(стр.
1901
—
14
)
Pespeni
MH
,
Garfield
DA
,
Manier
Mk
,
Palumbi
SR
.
Полногеномные полиморфизмы обнаруживают неожиданные цели естественного отбора
279
(стр.
1412
—
20
)
Песни
MH
,
Oliver
TA
,
Manier
MK
,
Palumbi
SR
.
Анализ тайлинга сайтов рестрикции: точное обнаружение и количественное генотипирование полиморфизмов всего генома с использованием массивов нуклеотидов
11
стр.
Р44
Песпени
MH
,
Палумби
SR
.
Сигналы отбора в аномальных локусах у широко расселенных видов в градиенте окружающей среды
22
(стр.
3580
—
97
)
Песни
MH
,
Сэнфорд
0004E
,
Gaylord
B
,
Hill
TM
,
Hosfelt
JD
,
Jaris
HK
,
LaVigne
M
,
Lenz
EA
,
Russell
AD
,
Young
MK
, 9007 и др.Эволюционные изменения во время экспериментального закисления океана
,
Proc Natl Acad Sci USA
,
2013b
, vol.
110
(pg.
6937
—
42
)
Pistevos
JCA
,
Calosi
P
,
Widdicombe
S
,
Bishop
JDD
.
Будут ли различия среди генетических особей влиять на реакцию видов на глобальное изменение климата?
,
Ойкос
,
2011
, том.
120
(стр.
675
—
89
)
Pörtner
H-O 90.0 90.
Экосистемные эффекты закисления океана во время его потепления: взгляд физиолога
373
(стр.
203
—
17
)
Pörtner
HO
,
Langenbuch
M
,
Michaelidis
B
.
Синергетическое воздействие экстремальных температур, гипоксии и повышения концентрации CO 2 на морских животных: от истории Земли к глобальным изменениям
110
стр.
C09S10
Проссер
CL
.
Адаптационная биология: молекулы для организмов
,
1986
Нью -Йорк (Нью -Йорк)
Wiley
R Основная команда разработки
R: Язы
Австрия: R Foundation for Statistical Computing
Ridgwell
A
,
Schmidt
D
,
Turley
C
,
Brownlee
C
,
Maldonado
MT
,
Tortell
P
,
Young
JR
.
От лабораторных манипуляций к моделям системы Земля: масштабирование последствий кальцификации подкисления океана
,
Биогеонауки
,
2009
, том.
6
(стр.
2611
—
23
)
Rogers-Bennett
7 L4
2 Лоуренс
ДМ
.
Экология Strongylocentrotus franciscanus и Strongylocentrotus purpuratus
,
Съедобные морские урчины: биология и экология
,
2007
Amsterm
,
2007
Amsterm9
004
(стр.
393
—
425
)
Санфорд
E
,
Kelly
M
.
Местная адаптация у морских беспозвоночных
3
(стр.
509
—
35
)
Selkoe
K
,
Watson
J
,
White
C
,
Horin
TB
,
Iacchei
M
,
Mitarai
S
,
Siegel
DA
,
Гейнс
SD
,
Тунен
RJ
.
Преодоление хаоса генетической мозаичности: генетика морских ландшафтов выявляет экологические и океанографические факторы, определяющие генетические закономерности у трех видов рифов умеренного пояса
,
Мол Экол
,
2010
, том.
19
стр.
3708
Слаткин
М
.
Поток генов и географическая структура природных популяций
236
(стр.
787
—
92
)
Smith
PE
,
Эппли
RW
.
Первичная продукция и популяция анчоуса в Южно-Калифорнийском заливе: сравнение временных рядов
27
(стр.
1
—
17
)
Snyder
MA
,
Sloan
LC
,
.0002 Diffenbaugh
NS
,
Bell
JL
.
Будущие изменения климата и апвеллинг в Калифорнийском течении
,
Geophys Res Lett
,
2003
, vol.
30
Spicer
J
,
Widdicombe
S
,
Needham
H
,
Berge
J
H
,
H
,
.0004
.
Влияние CO 2 -Ацидифицированная морская вода внеклеточной кислоты -базы северного морского ежа Стронглоцентрот Dröebachiensis
,
J exp Mar Biol ECOL
,
1114
9000 2.
407
(стр.
19
—
25
)
Strathmann
MF4 MF.
В кн.: Размножение и развитие морских беспозвоночных северного побережья Тихого океана: данные и методы изучения яиц, зародышей и личинок
,
1987
Сиэтл (WA)
Университет Вашингтонской прессы
(стр.
511
—
34
)
Strathmann
R 9000 40074.
Продолжительность пелагического периода у иглокожих с питающимися личинками из северо-восточной части Тихого океана
34
(стр.
23
—
8
)
Stumpp
M
,
,
Melzner
F
,
Thorndyke
M
,
Dupont
S
.
CO 2 Индуцированное окисление морской воды влияет на развитие личинок морского ежа I: повышенная скорость метаболизма снижает возможности роста и вызывает задержку развития
,
Comp Biochem Physiol A Mol Integr Physiol
,
2011
, том.
160
(pg.
331
—
40
)
Sunday
JM
,
Crim
RN
,
Harley
CDG
,
Hart
МВт
.
Количественная оценка скорости эволюционной адаптации в ответ на закисление океана
,
PLoS One
,
2011
, том.
6
стр.
E22881
Swindell
W
,
Huebner
M
,
Weber
A
.
Паттерны пластической и адаптивной экспрессии генов, связанные с температурным стрессом у Arabidopsis thaliana
,
Наследственность
,
2007
, vol.
99
(pg.
143
—
50
)
Szászi
K
,
Grinstein
S
,
Orlowski
J
,
Kapus
А
.
Регуляция эпителиальной изоформы обменника Na+/H+ цитоскелетом
10
(стр.
265
—
72
)
Todgham
AE
,
Hofmann
GE
.
Транскриптомный ответ личинок морского ежа Строильный заводрский пурпурат на CO 2 -Осгивание морской воды
,
J Exp Biol
,
2009
, кет.
212
(стр.
2579
—
94
)
Vermeij
GJ
.
Географическое ограничение как руководство к причинам вымирания: случай холодных северных океанов в неогене
15
(стр.
335
—
56
)
Whitehead
A
,
Crawford
ДЛ
.
Изменчивость внутри и между видами в экспрессии генов: сырье для эволюции
15
(pg.
1197
—
211
)
Zaytsev
O
,
Cervantes-Duarte
R
,
Montante
O
,
Гальегос-Гарсия
А
.
Прибрежный апвеллинг на тихоокеанском шельфе полуострова Нижняя Калифорния
59
(стр.
489
—
502
)
Приложение 1
Методы
Сборник из Урчинса
THIRTO 1) и отправлен в морскую лабораторию Bodega. Для каждой популяции ежей засевали 0,5 М KCl в морской воде и смешивали гаметы 10 самок и 10 самцов для оплодотворения. Оплодотворенные эмбрионы из каждой популяции были разделены на две группы, и группы были помещены в два отдельных сосуда, один из которых поддерживался при температуре окружающей среды p CO 2 уровни (400 мкатм, современный глобальный средний атмосферный CO 2 ), а другой — повышенные уровни p CO 2 (900 мкатм, умеренный прогноз «интенсивного использования ископаемого топлива» на 2100 год) (Мосс и др., 2010). Культуры содержались с плотностью 0,66 личинок на миллилитр, что соответствует экологически значимой плотности (Strathmann, 1987). Сосуды с культурами держали в столах с морской водой при температуре 14°C. Морскую воду в сосудах постоянно перемешивали вибрирующими лопастями и каждые 2 дня 90% морской воды заменяли, удаляя воду обратной фильтрацией через сетку и заменяя отфильтрованной морской водой, предварительно уравновешенной до желаемого уровня p CO 2 . Начиная с 3-х суток после оплодотворения после каждого водообмена личинок кормили смесью водорослей ( Rhodomonas sp. и Dunaliella sp.) ad libitum . См. Песпени и др. (2013b) для получения полной информации о методах культивирования и химическом составе воды.
Популяционная геномика
Подводя итоги предыдущего исследования, можно сказать, что для каждой популяции мы отобрали примерно 1000 живых личинок на 1-й и 7-й день после оплодотворения для каждой обработки p CO 2 . Мы экстрагировали общую РНК из каждого пула личинок с помощью TRIzol и подготовили библиотеки кДНК для секвенирования с использованием набора TruSeq в соответствии с протоколом производителя (Illumina, Сан-Диего, Калифорния). Каждый образец был секвенирован на одной дорожке Illumina HiSeq, что дало примерно 80 миллионов прочтений по 50 пар оснований на образец (Microarray and Genomics Core Facility, Университет Юты, Солт-Лейк-Сити, Юта). Необработанные данные последовательности были обрезаны по качеству, а затем по длине с использованием программ набора инструментов FASTX (Gordon 2011) с минимальной оценкой качества по шкале Phred 30 и минимальной длиной 30 п.н. Мы сопоставили прочтения каждого образца со всеми предсказанными генами пурпурного морского ежа (загружены с www.spbase.org), используя программу Burrows-Wheeler Aligner (BWA) (Li and Durbin 2009).) и с использованием параметров по умолчанию, кроме n = 0,001, чтобы учесть высокую генетическую изменчивость пурпурных морских ежей (Консорциум по секвенированию генома морских ежей и др., 2006; Песпени и Палумби, 2013). SNP во всех образцах были обнаружены с использованием программ набора инструментов для анализа генома (GATK) (McKenna et al., 2010) и самых строгих критериев разработчика (http://www.broadinstitute.org/gatk/guide/). Оценки частоты аллелей для каждого образца личиночного пула были извлечены из полученных файлов .vcf путем деления количества чтений качества, сопоставленных с эталонным аллелем, на общее количество сопоставленных чтений качества в этой позиции с использованием пользовательского скрипта Python. (Доступен по запросу). Список этапов обработки данных, которые можно настроить, можно найти на сайте http://sfg.stanford.edu (De Wit et al. 2012).
Оценки частот аллелей у личинок, отобранных в день 1 в условиях окружающей среды CO 2 , были взяты для представления частот аллелей, присутствующих в диких популяциях. Оценки личинок, отобранных на 7-й день после культивирования при повышенном уровне CO 2 , были взяты для представления частот аллелей после лабораторного культивирования при высоком уровне CO 2 . Различия в частоте аллелей между этими двумя образцами «день × лечение» были взяты для представления изменений в частоте аллелей из-за смертности личинок. Альтернативными объяснениями различий в частоте аллелей могут быть (1) различия в размере личинок и, следовательно, в обилии транскриптов среди образцов (однако мы измерили минимальные различия в размерах между обработками, поэтому размер личинок вряд ли повлияет на оценки) или (2 ) различия в аллель-специфической экспрессии, но мы не нашли доказательств этого в данных (см. Pespeni et al. (2013b) для анализа и обсуждения).
Сбор данных о рН окружающей среды
Мы провели высокочастотные измерения локального рН с использованием автономного датчика рН, установленного на каждом участке исследования (Ф. Чан и др., в процессе подготовки). Датчик pH был установлен в приливных участках, где собирались взрослые морские ежи, чтобы наиболее точно регистрировать pH, с которым сталкиваются ежи в дикой природе (за исключением Алегрии, Калифорния, 34,47 ° с.ш., где записи с ближайшего доступного датчика в Ломпоке, Калифорния , 34,72° с.ш.). Датчик pH представляет собой модифицированную версию датчика SeaFET (Martz et al. 2010), в котором используется pH-зонд Honeywell DuraFET с ионно-чувствительным полевым транзистором, встроенный регистратор данных и блок питания (Evans et al. 2013). Измерения проводились каждые 10 минут в течение сезона апвеллинга с апреля по сентябрь 2011 года в керне. Количество времени, проведенное ниже pH 7,8, было рассчитано для каждого участка, чтобы отразить серьезность низкого pH, наблюдаемого на каждом участке. Участки были ранжированы с 1 по 6 в зависимости от времени, проведенного при pH ниже 7,8, при этом 1 провел больше всего времени при низком pH (таблица 1).
Тест на корреляцию между частотой аллеля и локальным рН
Для каждого SNP мы проверили корреляцию между частотой аллеля и ранговым локальным рН в шести местах исследования. Мы использовали частичные тесты Mantel, реализованные в среде программирования R (R Development Core Team 2009), с использованием функции «mantel.partial» в веганской библиотеке (Oksanen et al. 2008), контролирующей географическое расстояние. P -значения определялись путем случайной перестановки данных и пересчета корреляции 1000 раз. Перед проверкой корреляции мы стандартизировали дисперсию для каждой переменной, разделив ее на среднее значение и вычитая одно стандартное отклонение в соответствии с процедурой Selkoe et al. (2010).
Тест на взаимосвязь между функцией гена и корреляцией с локальным pH
Мы проверили неслучайную концентрацию SNP с высокой корреляцией с локальными условиями pH в определенных функциональных классах генов, подход, называемый анализом функционального обогащения. Мы охарактеризовали каждый ген, используя идентификаторы UniProt (Bairoch et al. 2009), затем категории биологических процессов Gene Ontology (GO) (Ashburner et al. 2000). Мы также включили список генов биоминерализации, генов, важных для роста скелета морских ежей (как в Pespeni et al. 2013a, 2013b). Чтобы уменьшить шум и сосредоточиться на более интересных паттернах генома, мы ограничили анализ функциональными классами, содержащими от 10 до 100 членов генов, исключая классы, которые были слишком узкими или широкими, соответственно. Было 1307 функциональных категорий, которые содержали от 10 до 100 генов-членов. Для каждой функциональной категории мы проверили корреляцию между принадлежностью к этой категории и оценкой гена, используя повторную выборку оценки гена, реализованную в ErmineJ (Lee et al. 2005). Для оценок генов мы использовали 9 логарифмических преобразований.0014 P — значение для каждого SNP, рассчитанное в частичном тесте Мантеля для корреляции между частотой аллеля и ранговым pH, описанным выше. Когда было несколько SNP на ген, мы использовали средний балл. Статистическая значимость определялась по 10 000 перестановок, и скорректированные значения P рассчитывались с использованием подхода Бенджамини-Хохберга (Бенджамини и Хохберг, 1995). Чтобы определить, были ли определенные функциональные категории чрезмерно представлены среди тех, у кого P <0,01, мы сравнили наблюдаемые частоты с ожидаемыми частотами выше и ниже порогового значения 9.0014 P — значение с использованием точного критерия Фишера.
Идентификация лучших генов-кандидатов и сигналов селекции
Мы идентифицировали главные гены-кандидаты для реакции на закисление путем изучения 318 SNP с корреляцией с локальным pH ( P < 0,01). Эти SNP продемонстрировали общие клинальные паттерны в отношении рангового времени, проведенного при pH <7,8 для каждого сайта. Мы сравнили частоты аллелей, рассчитанные по клинальным аллелям в дикой природе (день 1, 400 мкАТм), с частотами аллелей через 7 дней при низком pH (день 7; 9 дней). 00 мкатм). Предполагая, что эти аллели адаптированы к низкому рН, мы искали минимальные изменения частоты аллелей при экспериментальном закислении в популяциях, предположительно адаптированных к низкому рН (FC и ST), и более значительные изменения в популяциях с высоким рН (AL и TP). ). Если бы все популяции отреагировали на экспериментальное подкисление к 7-му дню, можно было бы ожидать низкой изменчивости частот аллелей среди популяций на 7-й день для личинок при 900 мкатм. Чтобы определить лучших кандидатов на реакцию на подкисление, мы сделали эти расчеты, как описано, и соответственно ранжировали локусы. Мы также визуально изучили паттерны во всех 318 локусах, написав сценарий для циклического просмотра и построения всех 318 локусов. На основе этих критериев были отобраны семь лучших кандидатов, хотя все 318 локусов с частотами аллелей для каждой популяции можно найти в дополнительной таблице S1.
Если бы экспериментальная селекция была нацелена на те же аллели, которые были адаптированы к местным условиям pH в дикой природе, мы могли бы ожидать, что степень генетических изменений в ответ на экспериментальное закисление (низкий pH, высокий уровень CO 2 ) будет наибольшей в популяциях, которые обычно имеют высокий pH в природе и более низкий в популяциях, которые обычно имеют низкий pH в природе. Чтобы проверить это, мы рассчитали среднюю разницу в частоте аллелей между личинками на 1-й день (400 мкатм) и личинками на 7-й день (900 мкатм) в наборах из двух популяций с высшим и низшим ранговыми условиями рН (FC и ST, AL и TP соответственно). Мы сравнили два распределения ответов, используя критерий Колмогорова-Смирнова (K-S) (функция ks.test в R), и сравнили эти средние ответы в условиях низкого pH, используя t -критерий, предсказывающий, что популяции с высоким рангом проявят наибольший отклик. (функция t.test в R).
Примечания автора
Из симпозиума «Понимание фенотипов первого порядка: транскриптомика для новых моделей», представленного на ежегодном собрании Общества интегративной и сравнительной биологии 3–7 января 2013 г. в Сан-Франциско, Калифорния.
© Автор, 2013 г. Опубликовано Oxford University Press от имени Общества интегративной и сравнительной биологии. Все права защищены. Для получения разрешений обращайтесь по электронной почте: journals. [email protected].
© Автор, 2013 г. Опубликовано Oxford University Press от имени Общества интегративной и сравнительной биологии. Все права защищены. Для получения разрешений обращайтесь по электронной почте: [email protected].
Раздел выпуска:
Понимание фенотипов первого порядка: транскриптомика для новых моделей
Скачать все слайды
Дополнительные данные
Дополнительные данные
Дополнительные данные — zip-файл
Реклама
Цитаты
Альтметрика
Дополнительная информация о метриках
Оповещения по электронной почте
Оповещение об активности статьи
Предварительные уведомления о статьях
Оповещение о новой проблеме
Получайте эксклюзивные предложения и обновления от Oxford Academic
Ссылки на статьи по телефону
Последний
Самые читаемые
Самые цитируемые
Экоиммунология: что нетрадиционные организмы говорят нам через два десятилетия
Биотические взаимодействия и будущее рыб на коралловых рифах: важность подходов, основанных на признаках
Потепление ускоряет начало реакции на молекулярный стресс и увеличивает смертность личинок атлантической трески
Шаг вперед: функциональное разнообразие и новые темы медленной локомоции у позвоночных
Лучшие практики биоинспирированного дизайна: ключевые темы и проблемы
Реклама
Реакция вашего организма на стресс
Обзор
Стресс — обычное явление. Хотя вы не можете удалить из своей жизни все факторы стресса, можно управлять стрессом и поддерживать свое здоровье. Это важно, потому что стресс может вызвать умственную усталость, раздражительность и бессонницу.
Но даже если вы знаете о физических последствиях стресса, вы можете не знать о различных стадиях стресса, известных как общий адаптационный синдром (ОАС). Когда вы понимаете разные стадии стресса и то, как организм реагирует на эти стадии, вам будет легче выявить у себя признаки хронического стресса.
Подробнее: 20 эффектов стресса на организм »
ГАЗ представляет собой трехэтапный процесс, который описывает физиологические изменения, происходящие в организме в условиях стресса. Ганс Селье, врач и исследователь, выдвинул теорию ГАС. Во время эксперимента с лабораторными крысами в Университете Макгилла в Монреале он наблюдал ряд физиологических изменений у крыс после того, как они подверглись стрессовым событиям.
С помощью дополнительных исследований Селье пришел к выводу, что эти изменения были не изолированным случаем, а скорее типичной реакцией на стресс. Селье определил эти стадии как тревогу, сопротивление и истощение. Понимание этих различных реакций и того, как они соотносятся друг с другом, может помочь вам справиться со стрессом.
Подробнее: 10 простых способов снять стресс »
1. Стадия реакции тревоги
Стадия реакции тревоги относится к начальным симптомам, которые испытывает организм при стрессе. Возможно, вы знакомы с реакцией «бей или беги», которая является физиологической реакцией на стресс. Эта естественная реакция подготавливает вас либо к бегству, либо к защите в опасных ситуациях. Ваш сердечный ритм увеличивается, надпочечники выделяют кортизол (гормон стресса), и вы получаете заряд адреналина, который увеличивает энергию. Эта реакция «бей или беги» возникает на стадии реакции тревоги.
2. Стадия сопротивления
После первоначального шока от стрессового события и реакции «бей или беги» тело начинает восстанавливаться. Он высвобождает меньшее количество кортизола, и ваш сердечный ритм и кровяное давление начинают нормализоваться. Хотя ваше тело входит в эту фазу восстановления, некоторое время оно остается в состоянии повышенной готовности. Если вы преодолеваете стресс и ситуация больше не является проблемой, ваше тело продолжает восстанавливаться до тех пор, пока уровень гормонов, частота сердечных сокращений и кровяное давление не достигнут предстрессового состояния.
Некоторые стрессовые ситуации продолжаются длительное время. Если вы не избавитесь от стресса и ваше тело будет оставаться в состоянии повышенной готовности, оно в конечном итоге адаптируется и научится жить с более высоким уровнем стресса. На этом этапе тело претерпевает изменения, о которых вы не подозреваете, пытаясь справиться со стрессом.
Ваше тело продолжает выделять гормон стресса, и ваше кровяное давление остается повышенным. Вы можете думать, что хорошо справляетесь со стрессом, но физическая реакция вашего тела говорит о другом. Если стадия резистентности продолжается слишком долго без пауз, чтобы компенсировать последствия стресса, это может привести к стадии истощения.
К признакам стадии резистентности относятся:
- раздражительность
- разочарование
- плохая концентрация
3. Стадия истощения
Эта стадия является результатом длительного или хронического стресса. Борьба со стрессом в течение длительного времени может истощить ваши физические, эмоциональные и умственные ресурсы до такой степени, что у вашего тела больше не будет сил бороться со стрессом. Вы можете сдаться или почувствовать, что ваше положение безнадежно. К признакам истощения относятся:
- усталость
- выгорание
- депрессия
- тревога
- снижение стрессоустойчивости
Физические эффекты этой стадии также ослабляют вашу иммунную систему и подвергают вас риску заболеваний, связанных со стрессом.
ГАЗ может возникнуть при любом типе стресса. К стрессовым событиям относятся:
- потеря работы
- проблемы со здоровьем
- финансовые проблемы
- распад семьи
- травма
Но, несмотря на то, что стресс неприятен, положительная сторона заключается в том, что GAS улучшает реакцию организма на стрессоры, особенно на стадии тревоги.
Реакция «бей или беги», возникающая на стадии тревоги, предназначена для вашей защиты. Более высокий уровень гормонов на этом этапе приносит вам пользу. Это дает вам больше энергии и улучшает вашу концентрацию, чтобы вы могли сосредоточиться и справиться с ситуацией. Когда стресс кратковременный или кратковременный, стадия тревоги не опасна.
При длительном стрессе это не так. Чем дольше вы справляетесь со стрессом, тем вреднее он наносит вашему здоровью. Вы также не хотите оставаться на стадии сопротивления слишком долго и рискуете войти в стадию истощения. Когда вы находитесь на стадии истощения, продолжительный стресс повышает риск хронического высокого кровяного давления, инсульта, сердечных заболеваний и депрессии. У вас также более высокий риск инфекций и рака из-за более слабой иммунной системы.
Поскольку устранить все факторы стресса невозможно, важно найти способы справиться со стрессом. Знание признаков и стадий стресса может помочь вам предпринять соответствующие шаги, чтобы контролировать уровень стресса и снизить риск осложнений.
Для каждой популяции ежей засевали 0,5 М KCl в морской воде и смешивали гаметы 10 самок и 10 самцов для оплодотворения. Оплодотворенные эмбрионы из каждой популяции были разделены на две группы, и группы были помещены в два отдельных сосуда, один из которых поддерживался при температуре окружающей среды p CO 2 уровни (400 мкатм, современный глобальный средний атмосферный CO 2 ), а другой — повышенные уровни p CO 2 (900 мкатм, умеренный прогноз «интенсивного использования ископаемого топлива» на 2100 год) (Мосс и др., 2010). Культуры содержались с плотностью 0,66 личинок на миллилитр, что соответствует экологически значимой плотности (Strathmann, 1987). Сосуды с культурами держали в столах с морской водой при температуре 14°C. Морскую воду в сосудах постоянно перемешивали вибрирующими лопастями и каждые 2 дня 90% морской воды заменяли, удаляя воду обратной фильтрацией через сетку и заменяя отфильтрованной морской водой, предварительно уравновешенной до желаемого уровня p CO 2 . Начиная с 3-х суток после оплодотворения после каждого водообмена личинок кормили смесью водорослей ( Rhodomonas sp. и Dunaliella sp.) ad libitum . См. Песпени и др. (2013b) для получения полной информации о методах культивирования и химическом составе воды. Необработанные данные последовательности были обрезаны по качеству, а затем по длине с использованием программ набора инструментов FASTX (Gordon 2011) с минимальной оценкой качества по шкале Phred 30 и минимальной длиной 30 п.н. Мы сопоставили прочтения каждого образца со всеми предсказанными генами пурпурного морского ежа (загружены с www.spbase.org), используя программу Burrows-Wheeler Aligner (BWA) (Li and Durbin 2009).) и с использованием параметров по умолчанию, кроме n = 0,001, чтобы учесть высокую генетическую изменчивость пурпурных морских ежей (Консорциум по секвенированию генома морских ежей и др., 2006; Песпени и Палумби, 2013). SNP во всех образцах были обнаружены с использованием программ набора инструментов для анализа генома (GATK) (McKenna et al., 2010) и самых строгих критериев разработчика (http://www.broadinstitute.org/gatk/guide/). Оценки частоты аллелей для каждого образца личиночного пула были извлечены из полученных файлов .vcf путем деления количества чтений качества, сопоставленных с эталонным аллелем, на общее количество сопоставленных чтений качества в этой позиции с использованием пользовательского скрипта Python. (Доступен по запросу). Список этапов обработки данных, которые можно настроить, можно найти на сайте http://sfg.stanford.edu (De Wit et al. 2012). Pespeni et al. (2013b) для анализа и обсуждения). Участки были ранжированы с 1 по 6 в зависимости от времени, проведенного при pH ниже 7,8, при этом 1 провел больше всего времени при низком pH (таблица 1). Мы охарактеризовали каждый ген, используя идентификаторы UniProt (Bairoch et al. 2009), затем категории биологических процессов Gene Ontology (GO) (Ashburner et al. 2000). Мы также включили список генов биоминерализации, генов, важных для роста скелета морских ежей (как в Pespeni et al. 2013a, 2013b). Чтобы уменьшить шум и сосредоточиться на более интересных паттернах генома, мы ограничили анализ функциональными классами, содержащими от 10 до 100 членов генов, исключая классы, которые были слишком узкими или широкими, соответственно. Было 1307 функциональных категорий, которые содержали от 10 до 100 генов-членов. Для каждой функциональной категории мы проверили корреляцию между принадлежностью к этой категории и оценкой гена, используя повторную выборку оценки гена, реализованную в ErmineJ (Lee et al. 2005). Для оценок генов мы использовали 9 логарифмических преобразований.0014 P — значение для каждого SNP, рассчитанное в частичном тесте Мантеля для корреляции между частотой аллеля и ранговым pH, описанным выше. Когда было несколько SNP на ген, мы использовали средний балл. Статистическая значимость определялась по 10 000 перестановок, и скорректированные значения P рассчитывались с использованием подхода Бенджамини-Хохберга (Бенджамини и Хохберг, 1995). Чтобы определить, были ли определенные функциональные категории чрезмерно представлены среди тех, у кого P <0,01, мы сравнили наблюдаемые частоты с ожидаемыми частотами выше и ниже порогового значения 9.0014 P — значение с использованием точного критерия Фишера. 00 мкатм). Предполагая, что эти аллели адаптированы к низкому рН, мы искали минимальные изменения частоты аллелей при экспериментальном закислении в популяциях, предположительно адаптированных к низкому рН (FC и ST), и более значительные изменения в популяциях с высоким рН (AL и TP). ). Если бы все популяции отреагировали на экспериментальное подкисление к 7-му дню, можно было бы ожидать низкой изменчивости частот аллелей среди популяций на 7-й день для личинок при 900 мкатм. Чтобы определить лучших кандидатов на реакцию на подкисление, мы сделали эти расчеты, как описано, и соответственно ранжировали локусы. Мы также визуально изучили паттерны во всех 318 локусах, написав сценарий для циклического просмотра и построения всех 318 локусов. На основе этих критериев были отобраны семь лучших кандидатов, хотя все 318 локусов с частотами аллелей для каждой популяции можно найти в дополнительной таблице S1. Чтобы проверить это, мы рассчитали среднюю разницу в частоте аллелей между личинками на 1-й день (400 мкатм) и личинками на 7-й день (900 мкатм) в наборах из двух популяций с высшим и низшим ранговыми условиями рН (FC и ST, AL и TP соответственно). Мы сравнили два распределения ответов, используя критерий Колмогорова-Смирнова (K-S) (функция ks.test в R), и сравнили эти средние ответы в условиях низкого pH, используя t -критерий, предсказывающий, что популяции с высоким рангом проявят наибольший отклик. (функция t.test в R). [email protected].Дополнительные данные
Последний
Самые читаемые
Самые цитируемые
Хотя вы не можете удалить из своей жизни все факторы стресса, можно управлять стрессом и поддерживать свое здоровье. Это важно, потому что стресс может вызвать умственную усталость, раздражительность и бессонницу. Селье определил эти стадии как тревогу, сопротивление и истощение. Понимание этих различных реакций и того, как они соотносятся друг с другом, может помочь вам справиться со стрессом. Хотя ваше тело входит в эту фазу восстановления, некоторое время оно остается в состоянии повышенной готовности. Если вы преодолеваете стресс и ситуация больше не является проблемой, ваше тело продолжает восстанавливаться до тех пор, пока уровень гормонов, частота сердечных сокращений и кровяное давление не достигнут предстрессового состояния.